Nouveau traitement du syndrome du côlon irritable par remodelage du système nerveux autonome:
Introduction
Le syndrome du côlon irritable (SCI), est classé comme une maladie intestinale fonctionnelle. Le remodelage du système nerveux autonome par rééducation limbique est une technique très sophistiquée permettant un traitement physiologique des maladies intestinales. En cas de «syndrome du côlon irritable pur», l’efficacité du traitement est désormais confirmée (des centaines de patients traités avec succès) et il en va de même pour le SCI associé à une colite non spécifique. Maintenant, nous commençons à traiter avec succès les patients atteints de Maladies Inflammatoires de l’Intestin (MII). Pour comprendre l’efficacité de cette technique, nous devons commencer à étudier l’étiologie et le mécanisme exact conduisant à ces anomalies.
Le Syndrome de Côlon Irritable est une maladie chronique du tractus gastro-intestinal inférieur qui affecte jusqu’à 15% des adultes dans le monde. Difficilement caractérisés par des anomalies structurelles, une infection ou des troubles métaboliques, les mécanismes sous-jacents du SCI sont restés flous pendant de nombreuses années, mais des recherches récentes ont conduit à une meilleure compréhension de cette pathologie. En conséquence, le SCI est souvent appelé côlon spastique, nerveux ou irritable. Sa caractéristique est une douleur ou une gêne abdominale associée à un changement de consistance et / ou de fréquence des selles.
La fréquence du Syndrome du Côlon Irritable dans une population donnée dépend, en partie, de l’origine ethnique et culturelle et des critères utilisés pour diagnostiquer la maladie. 8 à 20% des adultes dans le monde occidental rapportent des symptômes compatibles avec le SCI (environ 65% d’entre eux sont des femmes). L’Asie et l’Afrique ont des taux similaires à ceux du monde occidental. En Inde, le SCI est plus fréquent chez les hommes, bien qu’il soit possible que cela résulte de différences dans la déclaration des symptômes et l’utilisation des soins de santé entre les sexes.
Les symptômes du Syndrome du Côlon Irritable peuvent inclure des douleurs abdominales, une distension, des ballonnements, une indigestion et divers symptômes de défécation. Il existe trois sous-catégories de SCI, selon les principaux symptômes dont :
- Des douleurs associées à la diarrhée;
- Des douleurs associées à de la constipation;
- Des douleurs associées à de la diarrhée alternant avec de la constipation.
Les principales caractéristiques du syndrome comprennent la motilité, la sensation et le dysfonctionnement du système nerveux central. Un dysfonctionnement de la motilité peut se manifester par des spasmes musculaires; les contractions peuvent être très lentes ou rapides. Une sensibilité accrue aux stimuli provoque des douleurs et une gêne abdominale. La recherche estime que de nombreux patients atteints du Syndrome du Côlon Irritable ont des contractions coliques désorganisées et sensiblement plus intenses que les témoins normaux. Le SCI n’est pas un trouble psychiatrique, bien qu’il soit lié au stress émotionnel et social, qui peut affecter à la fois l’apparition et la gravité des symptômes. Les patients sujets au Syndrome du Côlon Irritable souffrent d’un taux disproportionnellement plus élevé de comorbidité avec d’autres troubles, tels que la fibromyalgie, la fatigue chronique, les douleurs pelviennes et les troubles psychiatriques (angoisse, dépression..) ainsi que les autres maladies dont l’étiologie reste inconnue. Nous sommes définitivement convaincus que le SCI est directement lié aux troubles de fonctionnement du système limbique en raison de la réponse thérapeutique définitive que nous avons obtenu avec les patients concernés.
Pour expliquer clairement comment le système est perturbé et comment il crée d’innombrables maladies actuellement considérées comme incurables alors que notre centre parvient à les traiter de manière curative et définitive, nous allons commencer par le stress et le traumatisme.
I. Définition scientifique du stress
Le stress est une réponse biologique et psychologique d’adaptation de l’organisme face à une situation perçue comme une menace, un défi ou une contrainte (appelée facteur de stress).
Scientifiquement, le stress correspond à l’ensemble des réactions neurobiologiques, hormonales, cognitives et comportementales mobilisées pour maintenir l’homéostasie (l’équilibre interne du corps).
- Le concept moderne du stress a été formalisé par Hans Selye au XXe siècle.
Sur le plan physiologique, le stress est une réaction adaptative non spécifique de l’organisme à toute demande qui lui est faite (Selye).
- En psychologie
Le stress est défini comme une transaction dynamique entre l’individu et son environnement, dans laquelle la situation est évaluée comme dépassant ses ressources et menaçant son bien-être.
Cette définition provient des travaux de Richard Lazarus et Folkman intitulés « Le modèle transactionnel ».
II. Mécanisme du circuit du stress

Fig 1 : Image illustrant une situation de stress
Sur l’image ci-dessus, on aperçoit clairement une situation de stress qui est déclenchée par deux types de facteurs:
- Le facteur de stress physique
Cela correspond à la douleur, provoquée par une lésion mécanique et capté par les nocicepteurs mécaniques et polymodaux ainsi que les terminaisons nerveuses libres. Sous la peau et dans les tissus, il existe des terminaisons nerveuses libres. Certaines ne réagissent qu’à la pression forte (nocicepteurs mécaniques), d’autres réagissent à tout : pression, chaleur extrême, brûlure chimique (polymodaux). Ce sont elles qui détectent l’agression. Une fois l’alarme déclenchée, le signal électrique doit voyager. Il emprunte deux types de « câbles électriques » :
- Les fibres Aδ: Grosses et rapides. Elles transmettent la douleur vive et immédiate.
- Les fibres C : Minces et lentes. Elles transmettent la douleur secondaire, sourde et diffuse (le lancement qui persiste après le choc).
- pour parvenir au corps cellulaire du neurone sensitif qui est dans le ganglion spinal (Cf. Fig. 2). Puis l’information douloureuse entre dans la moelle par la racine dorsale et la connexion se fait avec le 2ème nerf sensitif dans les Lames de Rexed 1,2,5 (I, II, V) qui sont des couches anatomiques de la moelle. Les couches numéro I, II et V sont les zones spécifiquement dédiées à la réception et au tri des signaux de la douleur.
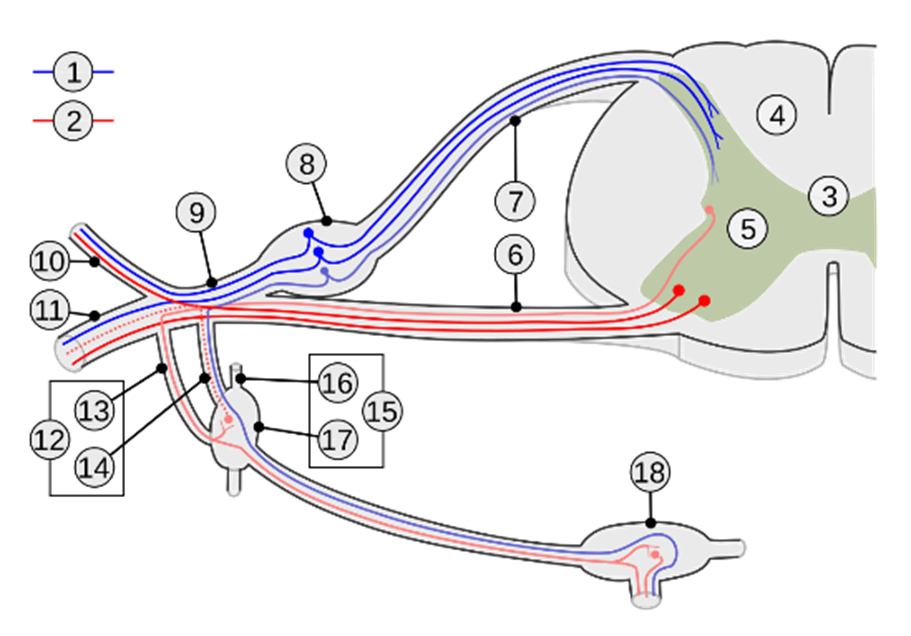 Fig 2 : Mécanisme du circuit du stress physique
Fig 2 : Mécanisme du circuit du stress physique
Le 2ème nerf sensitif, celui qui a reçu le relais dans les lames de Rexed de la moelle épinière, va se charger de faire monter le signal vers les centres supérieurs. Ainsi après décussation ou croisement, il passe du côté opposé de la moelle et puis remonte par la voie spinothalamique contra-latérale.
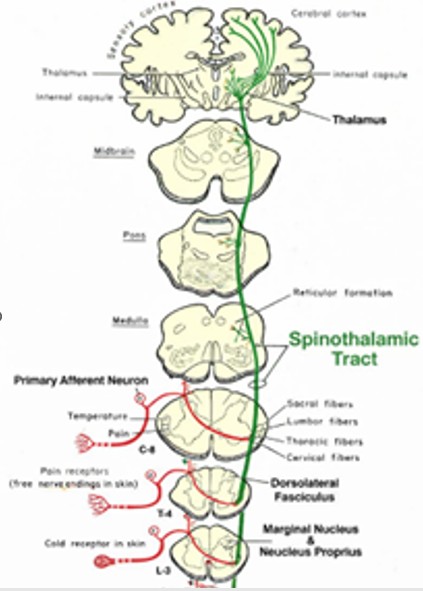 Fig. 3 : La voie spinothalamique
Fig. 3 : La voie spinothalamique
La voie spinothalamique dans le bulbe est située dans la partie latérale, en position dorso-latérale, proche du noyau spinal du trijumeau. Elle fait partie du système antérolatéral.
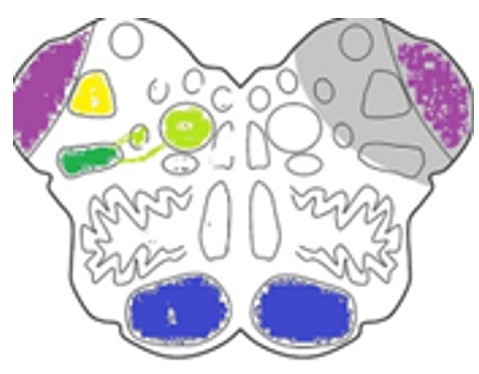 Fig. 4 : La voie spinothalamique et système antérolatéral dans le bulbe rachidien
Fig. 4 : La voie spinothalamique et système antérolatéral dans le bulbe rachidien
Les connexions au niveau bulbaire1. Collatérales vers la formation réticulée, participent à la voie spinoréticulaire, impliquée dans la composante émotionnelle de la douleur avec une projection ultérieure vers noyaux intralaminaires du thalamus
- Connexions avec le noyau spinal du trijumeau
Intégration de la douleur corporelle + la douleur faciale
- Participation à la voie spinoparabrachiale (indirecte), relais vers les centres autonomes
La voie spinothalamique dans le pont :
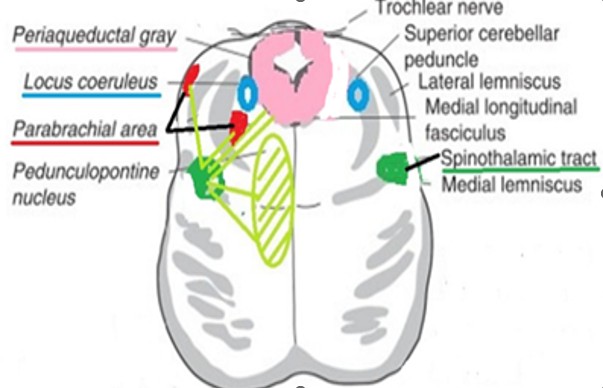
Fig 5 : La voie spinothalamique dans le pont
Les connexions au niveau du pont
- Formation réticulée pontique : modulation de la vigilance, participation aux réponses végétatives à la douleur
- Substance grise périaqueducale (via projections ascendantes), connexion indirecte, début des mécanismes de modulation descendante
3.Complexe para-brachial : Projection vers hypothalamus, réponse neurovégétative, composante affective
La voie spinothalamique dans le mésencéphale:
 Les connexions mésencéphaliques majeures
Les connexions mésencéphaliques majeures
- Substance grise périaqueducale (SGPA / PAG)
Centre majeur de modulation de la douleur, déclenche voie descendante inhibitrice, active noyau du raphé magnus → inhibition médullaire
- Colliculus supérieur (collatérales mineures), réflexes d’orientation vers stimulus nociceptif
- Formation réticulée mésencéphalique : Composante éveil / alerte.
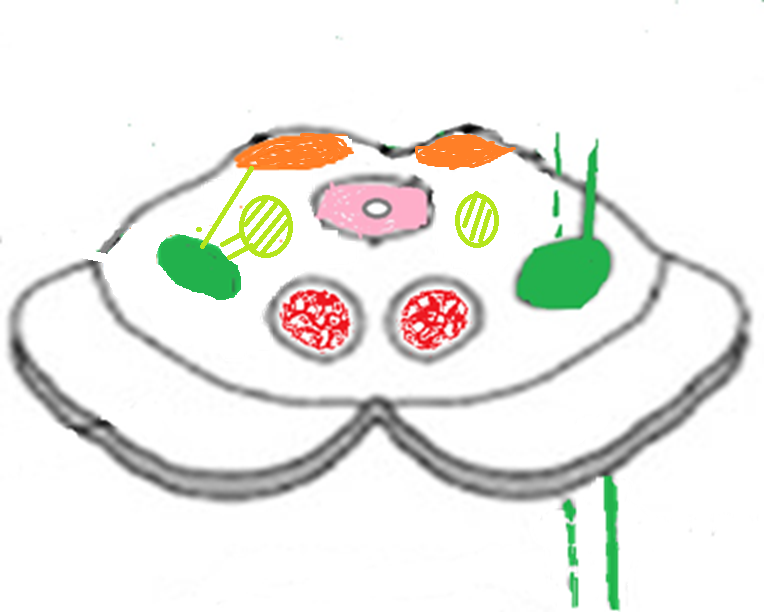
Thalamus (relais majeur)
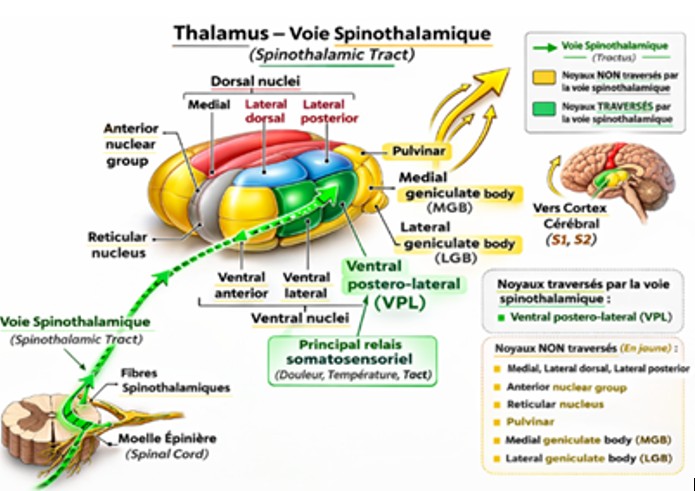 Fig. 6 Thalamus et voie spinothalamique
Fig. 6 Thalamus et voie spinothalamique
Noyaux thalamiques
VPL (ventro-postéro-latéral) → douleur du corps
Noyaux intralaminaires → composante émotionnelle
Au niveau du cortex cérébral (perception consciente), c’est le cortex somatosensoriel primaire qui reçoit l’information et détermine la localisation de la douleur et son Intensité.
- Autres régions activées
- Cortex somatosensoriel secondaire (S2)
- Insula → ressenti corporel
- Cortex cingulaire antérieur → souffrance / émotion
- Amygdale → peur, fuite
Le facteur de stress psychologique, la peur:
La peur est une émotion primaire de survie dont la fonction est de détecter un danger et de déclencher des réponses rapides (fuite, lutte, immobilisation). Chez la gazelle, elle est essentielle à la survie.
- Détection rapide du danger : Cette détection va se faire par les organes de sens : Visuel, auditif, olfactif et le toucher (somesthésique)
- Puis l’information est transmise au thalamus
- Et du thalamus vers l’amygdale,
Cette transmission se fait par deux voies différentes (selon Ledoux), la voie rapide est directe sans passer par le cerveau, mais elle est peu précise et est subconsciente (la confusion entre une corde et un serpent est possible) et la voie lente corticale qui passe d’abord au cortex sensoriel avant d’arriver à l’amygdale et qui est plus précise et détaillée (consciente).
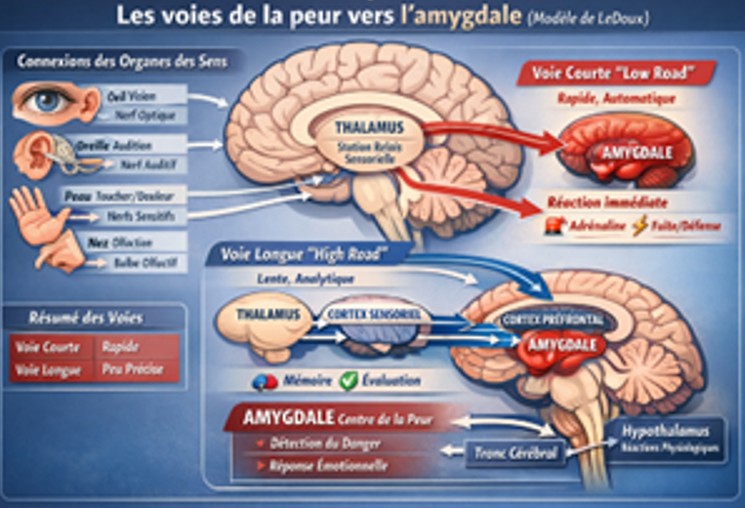
Fig. 7 Les voies de la peur vers l’amygdale
- Réponses de l’organisme au stress
- Réponse au facteur de stress physique
L’information arrive à l’hypothalamus (par les noyaux parabrachiaux et/ou par l’amygdale) , qui lance la procédure de fight and flight.
a) réponse autonome (immédiate) (Via l’hypothalamus et le tronc cérébral)
- Activation du système sympathique :
Tachycardie, Augmentation de la pression artérielle, Vasoconstriction des artères abdominales et périphériques, vasodilatation des artères musculaires, Mydriase, Bronchodilatation, Sudation, Inhibition de la motilité digestive et diminution temporaire de la production d’urine car le corps privilégie les fonctions vitales (cœur, muscles). Le flux sanguin vers les reins diminue → filtration réduite → moins d’urine produite à court terme. Parfois besoin urgent d’uriner (effet paradoxal). Cela correspond à la réaction « fight or flight ».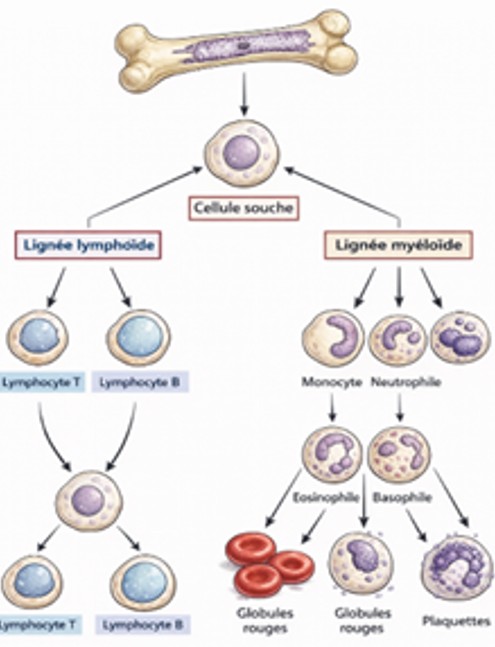
Fig.8 La stimulation des cellules souches
L’adrénaline libérée va agir sur tous les tissus, y compris sur la moelle osseuse. Les cellules souches sont stimulées pour devenir mûres et celles-ci sont mobilisées vers la circulation et incitées à déclencher l’inflammation car le système prévoit l’infection et par conséquence la défense (ce chapitre sera détaillé plus tard dans le cadre de stress chronique).
b) L’axe hypothalamo-hypophyso-surrénalien (HPA)
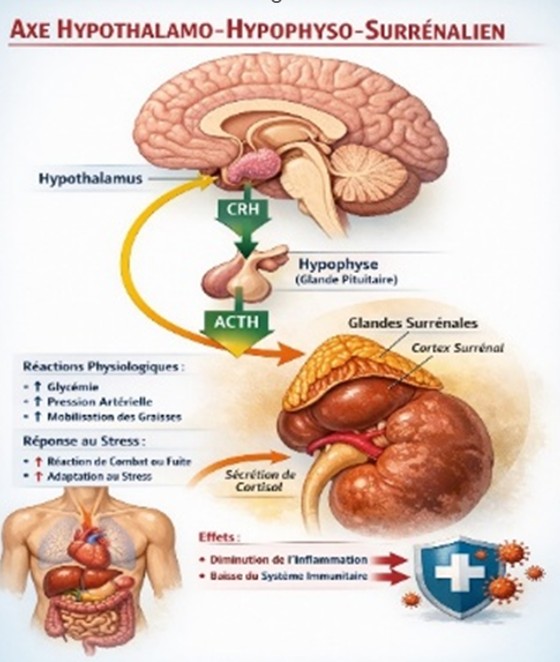
Fig 9. L’axe hypothalamo-hypophyso-surrénalien
L’axe HHS provoque la libération du cortisol qui est anti-inflammatoire dans la phase aiguë.
2) Réponse au facteur de stress psychologique (la peur)
La peur est une émotion primaire de survie dont la fonction est : détecter un danger et déclencher des réponses rapides (fuite, lutte, immobilisation). Chez la gazelle, elle est essentielle à la survie.
Détection rapide du danger : cette détection va se faire par les organes de sens : visuel, auditif, olfactif et le toucher (somesthésique).
Les informations arrivent au thalamus, qui agit comme un centre de tri.
Entre le thalamus et l’amygdale, il existe deux voies de transmission (modèle de LeDoux) :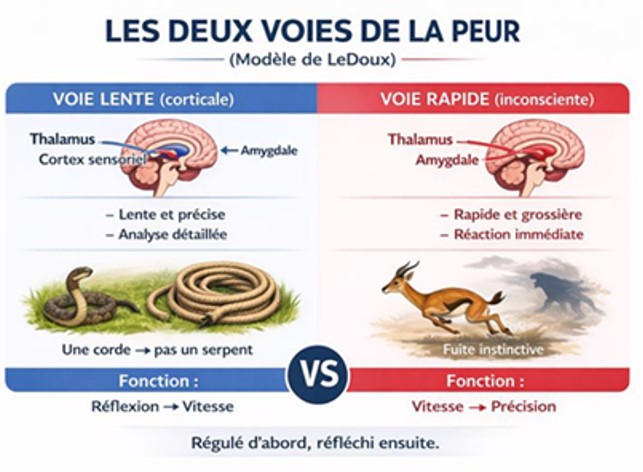
Fig 10. Les deux voies de transmission de la peur
- Voie rapide (subconsciente). Thalamus → Amygdale : Très rapide, Grossière, Permet une réaction immédiate (la fuite).Voie lente (corticale). Thalamus → Cortex sensoriel → Amygdale : Plus précise, plus lente, analyse détaillée.
- Réponses déclenchées par l’amygdale
1.1 Réponse autonome (immédiate)
Via l’hypothalamus et le tronc cérébral : activation du système sympathique avec tachycardie, augmentation de la pression artérielle, vasoconstriction périphérique, mydriase, bronchodilatation, sudation, inhibition digestive.
Cela correspond à la réaction « fight or flight ».
- a) Activation sympathique (globale), Voie :
- Amygdale → noyau central
- Projection vers hypothalamus latéral
- Activation des centres sympathiques bulbaires
- Descente vers la moelle thoraco-lombaire (T1–L2)
Tachycardie, voie précise : Amygdale → hypothalamus : Activation des neurones sympathiques préganglionnaires (T1–T5), Synapse dans les ganglions cervicaux, Fibres postganglionnaires → cœur.
Neurotransmetteur : noradrénaline, récepteur bêta.
- b) Les deux grands types de récepteurs impliqués
Vasoconstriction ou vasodilatation, voie : Amygdale → hypothalamus latéral, Activation sympathique médullaire, Fibres postganglionnaires → vaisseaux périphériques Neurotransmetteur : Noradrénaline
La noradrénaline agit principalement sur deux types de récepteurs :
- Récepteurs α1 (alpha-1) → vasoconstriction (artères du tube digestif)
α1 → protéine Gq → ↑ IP3 + Ca²⁺ et Ca²⁺ → contraction → vasoconstriction
- Récepteurs β2 (bêta-2) → vasodilatation (les artères des muscles squelettiques)
β2 → protéine Gs → ↑ AMPc inhibe la contraction → relaxation
➡️ Résultat : diminution du débit sanguin digestif
Mydriase
Amygdale → hypothalamus, Activation sympathique, Neurones préganglionnaires T1, Synapse dans le ganglion cervical supérieur, Fibres postganglionnaires → muscle dilatateur de l’iris. Récepteur : α₁
Sudation
Voie particulière (piège !) : Amygdale → hypothalamus, Activation sympathique, Fibres postganglionnaires → glandes sudoripares (ACH muscarinique)
Tableau ultra-synthèse
| Effet | Voie | NT final | Récepteur |
| Tachycardie | Sympathique T1–T5 | Noradrénaline | β₁ |
| Vasoconstriction | Sympathique | Noradrénaline | α₁ |
| Mydriase | T1 → ganglion cervical sup | Noradrénaline | α₁ |
| Sudation | Sympathique | ACh | Muscarinique |
Ce sont des noyaux du bulbe rachidien qui commandent le tonus sympathique en permanence et lors des réponses émotionnelles (via l’amygdale).
- c) RVLM – (Rostral Ventrolateral Medulla)
Localisation : Bulbe rachidien ventro-latéral
Rôle : Centre majeur du tonus sympathique, Maintient la pression artérielle, Active les neurones sympathiques médullaires.
Connexions :
- Reçoit des projections de : Hypothalamus et Amygdale (indirectement)
- Projette vers : Colonnes intermédio-latérales (T1–L2) de la moelle
- d) NTS – Noyau du tractus solitaire
Localisation : Bulbe dorsal
Rôle : Intègre les afférences viscérales, Baro- et chémorécepteurs, Module l’activité sympathique (plutôt régulation que commande directe).
- e) CVLM – Caudal Ventrolateral Medulla
Localisation : Bulbe ventro-latéral caudal
Rôle : Inhibiteur du RVLM, Participe au réflexe barorécepteur
Piège classique :
- CVLM ≠ centre sympathique actif
- rôle freinateur
- f) Formation réticulée bulbaire
Rôle : Ajustement global du tonus autonome, Lien émotions ↔ réponses végétatives.
1.2 Réponse hormonale
- Hypothalamus → hypophyse → surrénales
- Adrénaline + cortisol
1.3 Réponse comportementale
- Fuite explosive
- Hypervigilance
➡️ Peur = mécanisme normal, adaptatif et réversible
En résumé, ce stress, qui est un mécanisme physiologique, va aider cette gazelle à devenir plus courageuse et plus puissante, à tel point qu’elle va se tourner et va tuer le félin par un coup de corne dans le ventre. Et puis elle va aller se reposer dans un coin jusqu’à la guérison de ses blessures.
Après la guérison, deux situations complètement différentes vont se présenter :
- La Gazelle n’a pas été traumatisée : elle va avoir une vie normale
- La Gazelle a été traumatisée.
Dans ce dernier cas de figure, il va y avoir beaucoup de maladies et/ou de problèmes qui vont en découler.
Pour comprendre cela, nous allons voir ce système en détail :
- Le mécanisme des émotions
En 1937, dans son article scientifique intitulé “A Proposed Mechanism of Emotion”,Papez cherchait à expliquer le fonctionnement des émotions dans le cerveau. Il a proposé un circuit neuronal fermé, aujourd’hui appelé circuit de Papez, reliant plusieurs structures cérébrales.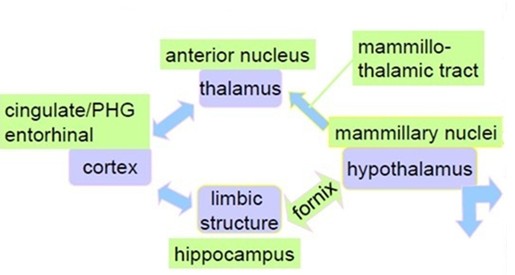
Fig 11. Circuit de Papez
Même si son modèle a été modifié ensuite (notamment avec le concept de système limbique), il reste une base historique majeure en neurosciences. Et puis on s’est rendu compte que ce système est beaucoup plus complexe et fait intervenir beaucoup de noyaux pour nous donner cette chance énorme d’avoir autant d’émotions et de plaisirs divers et variés.
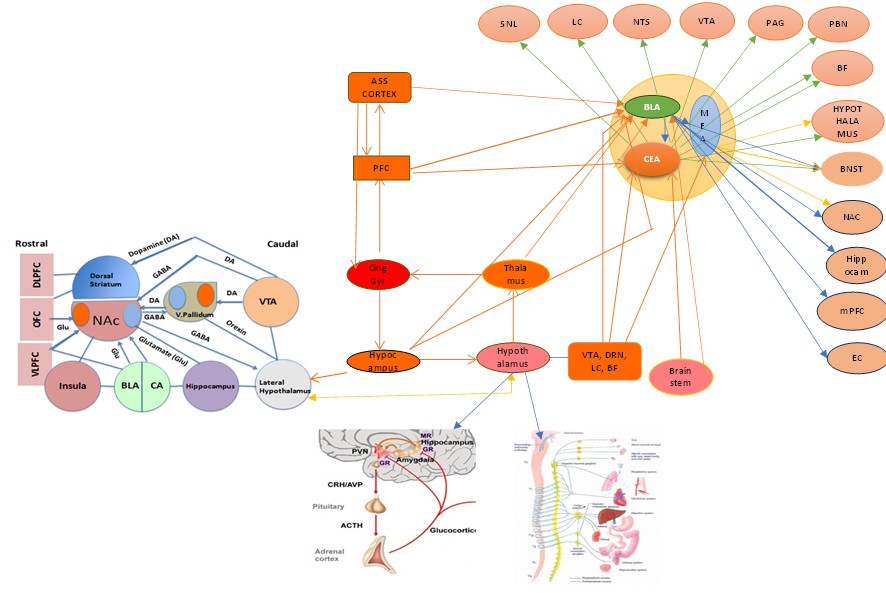
Fig 12. Le réseau de l’amygdale et de l’hippocampe dans le circuit de Papez
En résumé, l’information arrive à tous ces noyaux dans leur réseau respectif et l’amygdale avec l’hippocampe jouent un rôle important, l’un dans l’encodage de la valeur émotionnelle et l’autre dans l’enregistrement du contexte du lieu de traumatisme. Par exemple, pour cette gazelle, tout l’environnement va être enregistré subconsciemment ), mais le félin est enregistré consciemment. Puis le BLA stimule le BNST et celui-ci stimule le DMH et le PVN qui, à son tour, va agir sur le système sympathique et l’axe HHS (cf. plus haut).
L’amygdale est aussi en relation avec le cortex préfrontal qui a une action d’inhibition sur l’amygdale. Dans certaines conditions l’amygdale devient hyperactive et n’est plus sous contrôle du PFC (PreFrontal Cortex). Dans ces conditions, le stress est dominant et le patient est extrêmement anxieux.
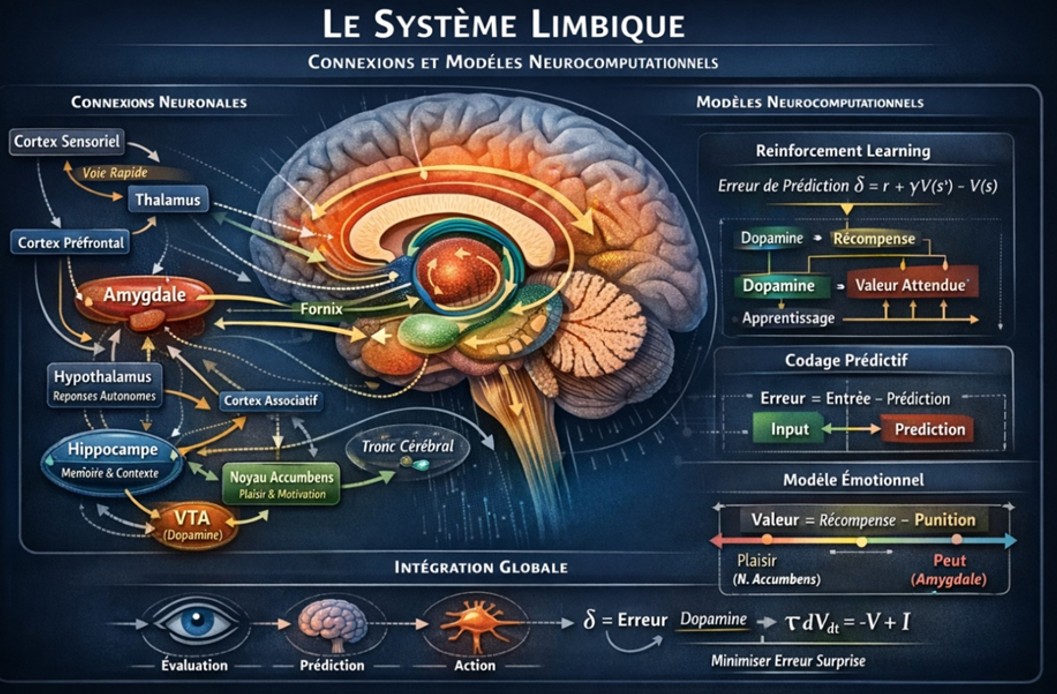
Fig 13. Le système limbique
2.1 Quand et comment la peur devient traumatique
Qu’est-ce qu’un traumatisme ? Un traumatisme survient quand la menace est extrême, la douleur et la peur sont intenses, il y a une impuissance, la situation déborde les capacités d’adaptation. Ici l’attaque avec griffes, et le sentiment de mort imminent peut être considéré potentiellement traumatique.
Dans ces situations il y a :
- Encodage anormal du souvenir traumatique
- L’amygdale est hyperactive et sur-encode la peur, y associe fortement les stimuli au danger.
- L’hippocampe est inhibé par le cortisol, ce qui entraîne un mauvais encodage du contexte (temps, lieu), un souvenir fragmenté et non daté.
Résultat : Le cerveau ne classe pas l’événement comme « passé ».
- Défaillance du cortex préfrontal :
Normalement, le cortex préfrontal inhibe l’amygdale, rassure en disant (« c’est fini »), calme toi …..
Sous traumatisme : activité diminuée, perte du contrôle top-down et l’amygdale reste hyperactive.
Manifestations possibles chez la gazelle :
- a) Hypervigilance
Réaction excessive au moindre bruit, fuite inappropriée.
- b) Réexpériences (équivalent animal)
Réactivation de la peur face à des stimuli similaires, Réactions automatiques non adaptées.
- c) Évitement : Abandon de zones pourtant sûres, Modification des comportements alimentaires.
- d) Sidération possible : Immobilité (freeze), Défaillance de la fuite.
Différence clé à retenir
| Peur normale | PTSD |
| Adaptative | Inadaptée |
| Temporaire | Persistante |
| Contextualisée | Décontextualisée |
| Réversible | Auto-entretenue |
Message fondamental
Le PTSD n’est pas un excès de peur, mais une mémoire de survie qui ne s’éteint pas.
Le traumatisme comme conditionnement pavlovien
Rappel du conditionnement pavlovien (classique)
Schéma simple : Stimulus neutre (SN) + stimulus aversif (SI) donne une réponse de peur. Après répétition ou intensité extrême, le stimulus neutre donne une réponse de peur.

Dans le trauma, une seule exposition peut suffire.
Le traumatisme = conditionnement pavlovien extrême
Stimulus inconditionnel (SI) : Attaque du prédateur, Douleur, Menace de mort
Déclenche automatiquement : Peur intense, Stress maximal, Réponses neurovégétatives.
Tous les éléments présents au moment du trauma peuvent devenir des déclencheurs ou Stimuli conditionnels (SC) : Odeur de l’herbe, Couleur du sol, Bruit du vent, Heure de la journée, Sensation corporelle (accélération cardiaque), Type de paysage (les collines, les cours d’eaux, herbes, fleurs, parfums des fleurs et des herbes…..), Les autres animaux, Les arbres, Bref une infinité d’éléments. Ainsi chaque élément peut devenir un déclencheur indépendant.
La BLA : le cœur émotionnel de la mémoire du stress
La BLA reçoit des informations sensorielles (thalamus + cortex), des signaux contextuels (hippocampe) et des signaux cognitifs (cortex préfrontal). Elle ne déclenche pas seulement la peur : elle associe un événement à une valeur émotionnelle forte (danger, stress, menace). Résultat : la BLA agit comme un amplificateur émotionnel des souvenirs.
Stress et encodage mnésique : ce qui se passe au moment du stress.
Lors d’un stress :
Activation neurochimique : libération de noradrénaline (locus coeruleus) et libération de cortisol (axe HPA) et la BLA est très riche en récepteurs :
- β-adrénergiques (noradrénaline)
- Glucocorticoïdes (cortisol)
Ces neuromodulateurs boostent l’activité synaptique dans la BLA.
Neuroplasticité dans la BLA : la clé de l’amplification
Plasticité synaptique (LTP)
Dans la BLA :
- ↑ Potentialisation à long terme (LTP)
- ↑ entrée de Ca²⁺ via NMDA
- Activation de CaMKII, PKA, MAPK
Les synapses associées au stress deviennent plus fortes , plus réactives, plus durables.
Un stimulus neutre associé au stress devient émotionnellement chargé.
Plasticité structurale
Le stress (surtout chronique) entraîne : ↑ densité des épines dendritiques dans la BLA ,↑ ramification dendritique, ↑ connectivité intra-amygdalienne.
La BLA devient hyperplastique . Contrairement à l’hippocampe (où le stress peut réduire laplasticité),.
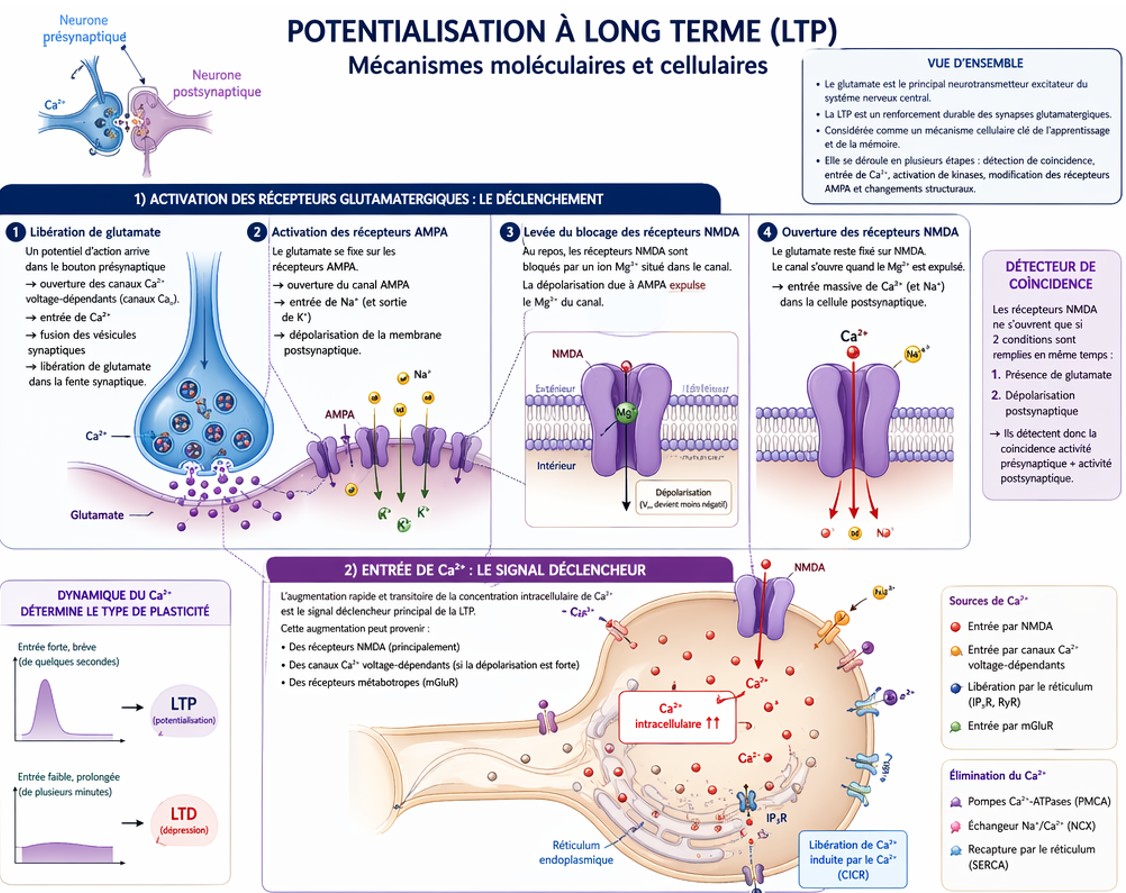
1) Potentialisation à long terme (LTP)
La LTP est une augmentation durable de l’efficacité de la transmission synaptique après une stimulation répétée et intense d’une synapse. Elle est surtout étudiée dans l’hippocampe (mémoire déclarative).
⚡ 2) ↑ Entrée de Ca²⁺ via les récepteurs NMDA
Le mécanisme clé débute au niveau des récepteurs NMDA :
- Le glutamate est libéré par le neurone présynaptique.
- Les récepteurs AMPA s’activent → dépolarisation postsynaptique.
- Cette dépolarisation enlève le blocage au Mg²⁺ des récepteurs NMDA.
- Les récepteurs NMDA s’ouvrent → entrée massive de Ca²⁺ dans la cellule.
Le Ca²⁺ est le signal déclencheur principal de la LTP.
3) Activation des kinases intracellulaires
L’augmentation du Ca²⁺ active plusieurs enzymes clés :
CaMKII (Calcium/Calmodulin-dependent protein kinase II)
- Activée directement par le complexe Ca²⁺/calmoduline
- Phosphoryle les récepteurs AMPA
- Augmente leur conductance
- Favorise l’insertion de nouveaux récepteurs AMPA dans la membrane
Ce qui entraîne un renforcement rapide de la synapse (phase précoce de la LTP).
PKA (Protein Kinase A)
- Activée via l’AMPc
- Participe à la stabilisation de la LTP
- Impliquée dans les changements transcriptionnels
MAPK (Mitogen-Activated Protein Kinase)
- Active des facteurs de transcription (ex : CREB)
- Permet la synthèse de nouvelles protéines
- Nécessaire pour la LTP tardive (L-LTP)
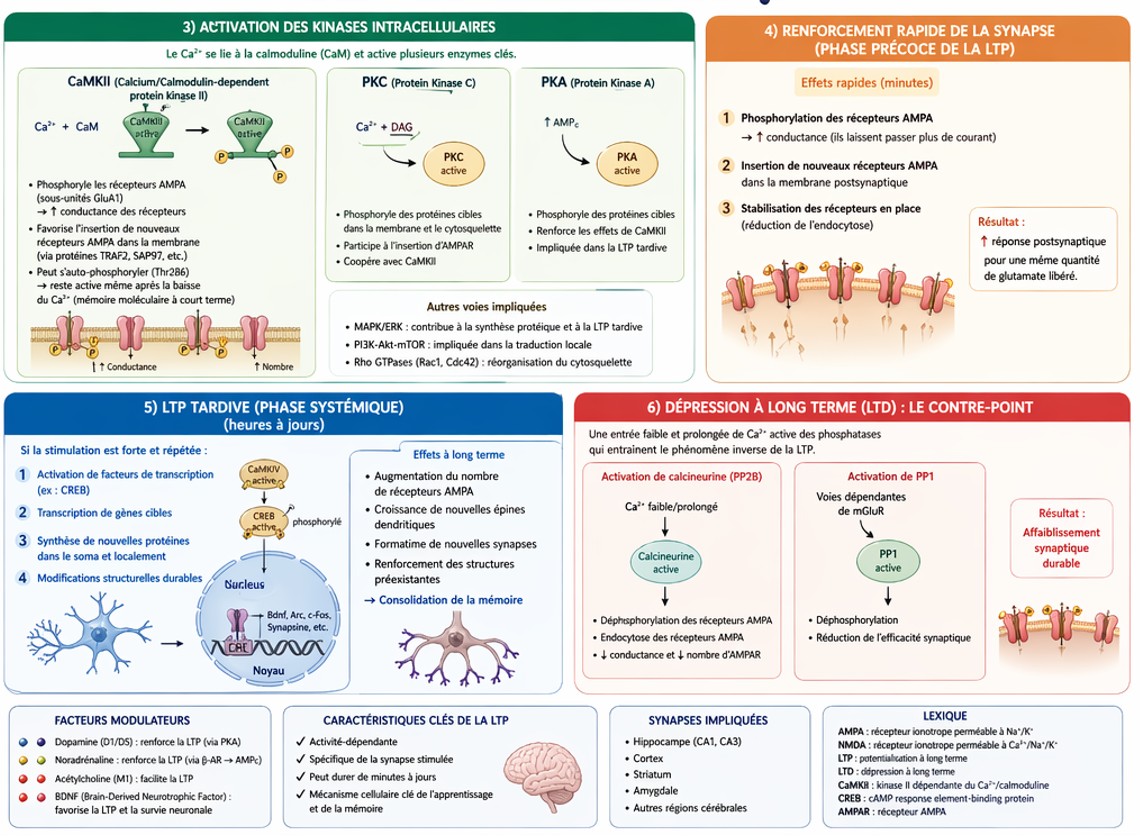
Résultat final
Augmentation du nombre et de l’efficacité des récepteurs AMPA, modifications structurelles (épines dendritiques), renforcement durable de la synapse
Résumé en chaîne logique
Stimulation intense →Activation AMPA →Activation NMDA →↑ Ca²⁺ → Activation CaMKII / PKA / MAPK →↑ Récepteurs AMPA + modifications structurelles → LTP (mémoire)
La relation entre le stress et l’inflammation:
Réponses immunitaires induites par le stress dans la périphérie et leurs mécanismes .
Mobilisation des leucocytes induite par le stress à partir de la moelle osseuse:
Des études sur des rongeurs, principalement avec un stress répété de défaite sociale, ont montré que le stress modifie le nombre, la distribution et les propriétés des leucocytes . En général, le stress chronique diminue les lymphocytes, tels que les cellules T et les cellules B, tandis qu’il augmente les cellules myéloïdes, telles que les neutrophiles et les monocytes, dans le sang et la moelle osseuse. Ces changements pourraient être dus à une altération de la différenciation et de la prolifération des leucocytes dans la moelle osseuse et de leur sortie vers la circulation. Ces ensembles de leucocytes ne se comportent pas de la même manière après la cessation du stress. L’augmentation des neutrophiles et la diminution des cellules B se maintiennent pendant plusieurs jours après le dernier stress, tandis que d’autres changements leucocytaires disparaissent. Le stress aigu pourrait induire des changements similaires dans les leucocytes que le stress chronique, car la diminution des lymphocytes et l’augmentation des cellules myéloïdes étaient perceptibles immédiatement après une exposition unique de 2 heures au stress de défaite sociale. Cependant, les effets du stress aigu peuvent être moins durables. Seule une augmentation des neutrophiles, mais pas une augmentation des monocytes, a été observée 12 heures après une exposition unique de 2 heures au stress de défaite sociale. Ces résultats indiquent que le stress chronique induit des changements leucocytaires plus durables que le stress aigu et que l’augmentation des neutrophiles dure plus longtemps que les changements dans d’autres sous-ensembles de leucocytes.
Rôles des systèmes autonome et neuroendocrinien
La mobilisation des leucocytes de la moelle osseuse induite par le stress est principalement médiée par les systèmes autonome et neuroendocrinien. L’activation du système nerveux sympathique lors du stress induit la libération de noradrénaline et d’adrénaline dans le sang et les tissus. Ces neurotransmetteurs activent les récepteurs adrénergiques sur les leucocytes et les cellules environnantes (par exemple, les cellules souches mésenchymateuses de la moelle osseuse), ce qui conduit à la mobilisation des leucocytes induite par le stress. L’activation sympathique tsf être suffisante pour ces effets, car le traitement systémique avec l’isoprénaline, un agoniste β-adrénergique non sélectif, diminue les lymphocytes et augmente les monocytes et les granulocytes dans la moelle o/*-Ces récepteurs adrénergiques pourraient favoriser la différenciation et la prolifération des cellules myéloïdes. Ils pourraient également réduire l’expression du ligand du motif C-X-C (CXCL) 12 sur les ostéoblastes et les cellules souches mésenchymateuses qui retiennent les cellules myéloïdes dans la moelle osseuse via le récepteur de chimiokine du motif C-X-C (CXCR) 4. De plus, le stress active l’axe HPA et augmente le taux de glucocorticoïdes dans le sang. Les glucocorticoïdes agissent directement sur les cellules myéloïdes et lymphoïdes pour supprimer les réponses inflammatoires. Ils favorisent également l’apoptose des cellules T, des macro phages et des cellules dendritiques, ce qui entraîne une inhibition des réponses immunitaires. Cependant, après un stress chronique, les monocytes et les macrophages développent une résistance aux glucocorticoïdes ou une sensibilité réduite aux glucocorticoïdes chez les souris et les humains, ce qui désinhibe les réponses inflammatoires. Parallèlement, les glucocorticoïdes favorisent les réponses immunitaires innées au stress en supprimant la voie CXCL12-CXCR4 qui retient les cellules myéloïdes dans la moelle osseuse et en augmentant la libération de cytokines par ces cellules, puisque la métyrapone, un inhibiteur de la synthèse des glucocorticoïdes, a empêché toutes ces réponses.
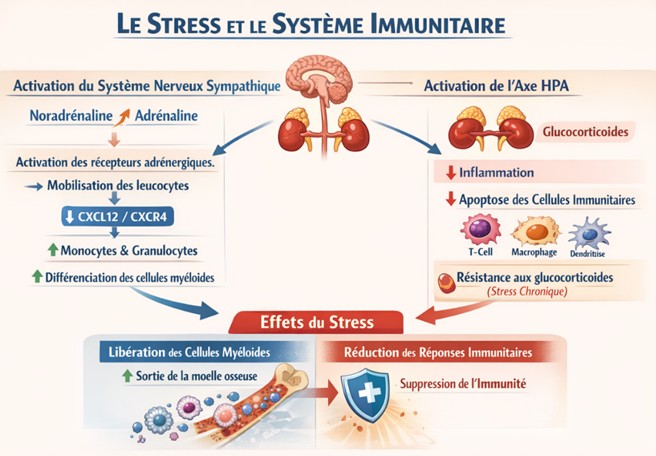
Rôles des cytokines et des chimiokines :
Le stress affecte l’expression de diverses cytokines et chimiokines dans la moelle osseuse et la circulation, contribuant probablement aux réponses immunitaires induites par le stress. Des cytokines distinctes mais qui se chevauchent régulent la prolifération et la mobilisation des neutrophiles et des monocytes. Le G-CSF(Granulocyte colony-stimulating factor) favorise généralement la prolifération et la mobilisation des neutrophiles, mais pas exclusivement. On pense que le G-CSF active le système nerveux sympathique localement dans la moelle osseuse, conduisant à la mobilisation des neutrophiles. CXCl 1*2 et leur récepteur CXCR2 stimulent également la mobilisation des neutrophiles. Le M-CSF(macrophage colony-stimulating factor) et le GM-CSF ( Granulocyte-Macrophage Colony-Stimulating Factor) augmentent la prolifération et mobilisation des monocytes, et la chimiokine à motif C-C 2 (CCL2) (chemokine ligand 2 , aussi appelée monocyte chemoattractant protein 1 (MCP1) et son récepteur C-C chimiokine de type 2 (CCR2) stimulent la mobilisation des monocytes. Le stress augmente les niveaux de G-CSF dans le sang. Comme l’IL-17 favorise la mobilisation des neutrophiles via la sécrétion de G-CSF, le stress de contention augmente les neutrophiles âgés via l’IL-17A, vraisemblablement dérivée des cellules T helper 17 (Th17) intestinales. Étant donné que cette mobilisation des neutrophiles dépend des bactéries filamenteuses segmentées dans l’intestin, l’IL-17A pourrait relier le microbiote intestinal et la réponse immunitaire innée au stress. En effet, le stress affecte le microbiote intestinal qui est associé à la susceptibilité au stress et l’influence. Bien que le mécanisme reste insaisissable, il a été démontré que le microbiote intestinal est régulé par de multiples voies, telles que les nerfs vagues, les glucocorticoïdes, les leucocytes et les cytokines sécrétées. Cependant, les effets du stress sur les taux d’IL-17 circulants varient selon les études, et il reste à établir si l’IL-17A intervient dans la mobilisation des neutrophiles sous stress. De plus, le stress augmente les taux de CXCL2 et de CCL2 dans le sang et le cerveau. Cette augmentation de CCL2 n’est pas essentielle à la mobilisation des monocytes induite par le stress dans la circulation puisque la suppression de CCR2 ne l’a pas affectée. Au contraire, l’augmentation de CCL2 dans le cerveau est cruciale pour l’infiltration des monocytes induite par le stress dans le cerveau.
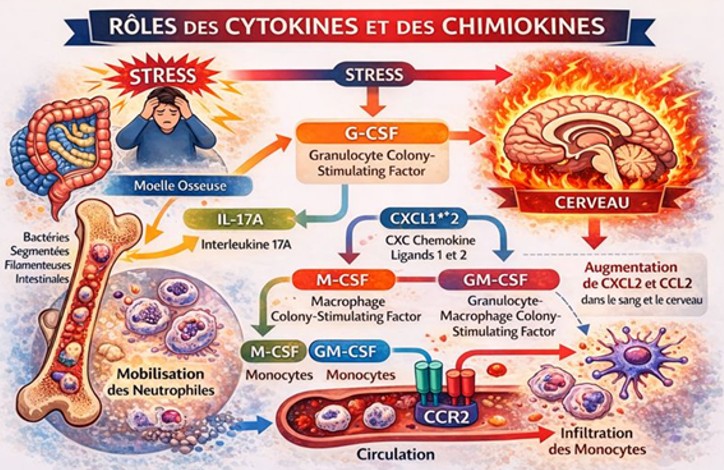
Les effets du stress sur le système immunitaire via plusieurs organes
Le stress peut affecter la rate et les ganglions lymphatiques via la projection du système nerveux sympathique, au moins en partie. Le stress aigu induirait une libération de noradrénaline par les terminaisons nerveuses sympathiques de la rate, qui agit sur les lymphocytes T choline-acétyltransférase-positifs (ChAT + ) via les adrénocepteurs β2. Ces lymphocytes T de la rate libèrent ensuite de l’acétylcholine et inhibent la production de cytokines pro-inflammatoires par les macrophages. Cette voie anti-inflammatoire est bénéfique dans divers modèles de maladies inflammatoires, notamment les lésions rénales aiguës ischémiques et peut être activée par les nerfs vagues connectés aux nerfs sympathiques spléniques. Le stress chronique, tel que les chocs électriques répétés au pied ou le stress de contention, augmente les cellules T CD4 + avec une glycolyse et une phosphorylation oxydative réduites ainsi qu’une fission mitochondriale dans la rate et les ganglions lymphatiques. Ce remodelage métabolique semble contribuer à l’anxiété élevée induite par le stress. Le stress chronique peut également augmenter les cellules plasmatiques spléniques et améliorer les réponses immunitaires dépendantes des anticorps en activant les régions cérébrales liées au stress, telles que le noyau central de l’amygdale et le noyau paraventriculaire de l’hypothalamus, ainsi que les nerfs spléniques. Cette amélioration des réponses humorales ne se produit que lorsque le stress est faible et ne provoque pas de libération de glucocorticoïdes, de sorte qu’elle ne se produit pas en cas de stress de contrainte accompagné de libération de glucocorticoïdes. Ainsi, l’immunité humorale est régulée différemment en fonction des conditions de stress. Le stress peut également affecter les organes périphériques non immunisés, car le stress aigu augmenterait la libération d’IL-6 par les adipocytes bruns par le biais d’actions sympathiques directes. Collectivement, les interactions entre plusieurs organes, notamment la moelle osseuse, la rate, l’intestin et le tissu adipeux brun, façonnent les réponses immunitaires innées et adaptatives induites par le stress d’une manière qui dépend des conditions de stress.
Rôles des cellules myéloïdes dérivées de la moelle osseuse.
Les réponses immunitaires induites par le stress décrites ci-dessus affectent les fonctions neuronales et contribuent aux changements comportementaux induits par le stress. Il existe de multiples barrières entre le cerveau et la périphérie, et les leucocytes existent rarement dans le parenchyme cérébral en bonne santé. Néanmoins, on pense que les cellules myéloïdes, telles que les monocytes, infiltrent les espaces périvasculaires du cerveau, en particulier en cas de stress chronique. Les monocytes circulants adhèrent aux cellules endothéliales vasculaires du cerveau et pénètrent dans l’espace périvasculaire. Parallèlement, une étude élégante sur la transplantation du crâne a démontré la voie directe des cellules myéloïdes de la moelle osseuse du crâne à la dure-mère, sautant l’entrée dans la circulation. Les cellules myéloïdes durales devraient encore pénétrer un autre type de barrière, l’arachnoïde et la pie-mère situés en dessous, pour atteindre le parenchyme cérébral. Le stress répété de défaite sociale augmente les monocytes périvasculaires d’une manière dépendante de CCR2. Il a été rapporté que CCL2, un ligand de CCR2, augmente dans la microglie après un stress répété de défaite sociale, suggérant que le CCL2 dérivé de la microglie recrute les monocytes périvasculaires via CCR2. Les souris dépourvues de CCR2 atténuent l’anxiété élevée induite par le stress répété de la défaite sociale et l’infiltration cérébrale concomitante de monocytes, suggérant le rôle anxiogène des monocytes transmissibles par le sang. Le stress répété de la défaite sociale mobilise également les neutrophiles dans la circulation, et cette augmentation des neutrophiles se maintient plus longtemps après la cessation du stress que celle des monocytes. Notamment, l’augmentation des neutrophiles est plus prononcée chez les souris BALB/c que chez les souris C57BL/6 N. Étant donné que les souris BALB/c sont plus sensibles au stress que les souris C57BL/6 N, l’augmentation des neutrophiles induite par le stress pourrait être un médiateur de la susceptibilité génétique au stress.
Rôles des cytokines circulantes
Les cellules myéloïdes mobilisées par le stress semblent affecter les fonctions neuronales via la sécrétion de cytokines. L’IL-6 est l’une de ces cytokines, car la transplantation de moelle osseuse de souris déficientes en IL-6 réduit le comportement dépressif. Notamment, une exposition unique de 10 minutes au stress de défaite sociale augmente l’IL-6 dans le sang avant que le comportement dépressif ne se développe. Cette augmentation d’IL-6 est plus importante chez les souris qui développeront éventuellement un comportement dépressif après le stress répété (c’est-à-dire les souris sensibles) que chez celles qui ne le développeront pas (c’est-à-dire les souris résilientes). Ainsi, la variabilité préexistante de la libération d’IL-6 induite par le stress par les leucocytes pourrait contribuer à la sensibilité au stress. L’IL-6 circulante semble s’infiltrer dans le parenchyme cérébral à travers la barrière hémato-encéphalique (BHE) car le stress répété de défaite sociale perturbe la BHE avec une réduction de la claudine-5 responsable des jonctions serrées dans les cellules endothéliales de la vascularisation cérébrale.
L’IL-1β et son récepteur, le récepteur de l’IL-1 de type I (IL-1RI), sont également impliqués dans les comportements dépressifs et anxieux induits par le stress. L’IL-1β dérivée des leucocytes semble cruciale pour l’anxiété élevée puisque la transplantation de moelle osseuse de souris dépourvues de caspase-1, une enzyme responsable de la maturation de l’IL-1β, a aboli ce changement de comportement (McKim et al., 2018). L’IL-1β dérivée des leucocytes pourrait agir sur l’IL-1RI dans la vascularisation cérébrale pour provoquer ses effets anxiogènes, comme le montre l’inactivation spécifique de l’IL-1RI dans les cellules endothéliales qui atténue l’anxiété élevée induite par le stress. Ces résultats illustrent que les cytokines dérivées des leucocytes affectent les fonctions neuronales par de multiples voies, directement ou indirectement (Fig. 2).
Rôles des lymphocytes T et autres cellules immunitaires
Comme décrit ci-dessus, le stress augmente également les lymphocytes T CD4 + avec un remodelage métabolique dans la rate et les ganglions lymphatiques. Les lymphocytes T CD4 + sont essentiels pour les comportements anxieux induits par le stress chronique, comme le démontrent la perte de ce comportement avec l’épuisement des lymphocytes T CD4 + médié par les anticorps ainsi que la suppression génétique de Rag1 (Fan et al., 2019). Transfert adoptif ont révélé que les lymphocytes T CD4 + naïfs de souris stressées sont suffisants pour restaurer le comportement anxieux chez les souris Rag1 knockout, suggérant des fonctions non conventionnelles des cellules immunitaires adaptatives. La fission mitochondriale des lymphocytes T CD4 + sous stress chronique pourrait être à l’origine d’un comportement anxieux induit par le stress puisque l’induction génétique de la fission mitochondriale dans ces cellules provoque un comportement anxieux même sans stress. L’effet anxiogène des lymphocytes T CD4 + avec fission mitochondriale dépend de la purine nucléoside phosphorylase 2 dans la voie de la xanthine oxydase, impliquant un métabolisme anormal de la purine dans cet effet comportemental. Les cellules immunitaires, y compris les macrophages et les lymphocytes résidents, existent autour des barrières entre le cerveau et la périphérie et pourraient être impliquées dans la neuroinflammation. Ces macrophages sont appelés macrophages associés au SNC (CAM), classés en macrophages périvasculaires, méningés et du plexus choroïde. Il a été récemment rapporté que les cellules T γδ méningées régulent le comportement anxieux via la signalisation IL-17A dans les neurones préfrontaux. Il reste à déterminer si le stress affecte les macrophages méningés et du plexus choroïde et les lymphocytes résidents et s’ils sont impliqués dans les dysfonctionnements neuronaux induits par le stress.
La relation entre le stress et l’intestin:
Le stress active deux grands systèmes :
A) Axe HPA (Hypothalamo-Hypophyso-Surrénalien)
a1) Le stress stimule l’hypothalamus qui secrète du CRH (Corticotropin-Releasing Hormone) , le CRH stimule l’hypophyse qui à son tour sécrète l’ACTH
a2) Puis l’ACTH agit sur les glandes surrénales pour libérer du cortisol dans la circulation
Le cortisol modifie l’immunité, le métabolisme et la barrière intestinale
B) Signaux périphériques intestinaux
CRH périphérique agit sur les neurones entériques, les cellules épithéliales et les cellules immunitaires y compris les mastocytes et l’activation des mastocytes provoque leur dégranulation autrement dit la libération de médiateurs inflammatoires tel que: TNF-α, IL-1β, tryptase, histamine
Ces molécules vont perturber directement la barrière intestinale
C) Système nerveux autonome
- ↓ tonus parasympathique (nerf vague)
- ↑ tonus sympathique
Le résultat de la phase d’activation initiale est l’inflammation
D) Signalisation intracellulaire (dans les entérocytes)
Cascade moléculaire :
Voie 1 :
- Activation de la protéine Gq/11
- Un ligand se fixe à un récepteur couplé aux protéines G (RCPG).
- Cela active la protéine Gq/11.
- Activation de la phospholipase C β (PLCβ)
- La sous-unité α de Gq active l’enzyme PLCβ.
- Hydrolyse du PIP₂
- La PLCβ clive le phosphatidylinositol 4,5-bisphosphate (PIP₂) en deux seconds messagers :
- IP₃ (inositol 1,4,5-trisphosphate)
- DAG (diacylglycérol)
- La PLCβ clive le phosphatidylinositol 4,5-bisphosphate (PIP₂) en deux seconds messagers :
- Action de l’IP₃
- L’IP₃ diffuse dans le cytosol et se fixe sur ses récepteurs situés sur le réticulum endoplasmique.
- Cela provoque la libération de Ca²⁺ intracellulaire.
- Conséquences
- L’augmentation du Ca²⁺ intracellulaire active diverses protéines dépendantes du calcium (calmoduline, MLCK, etc.).
- Le DAG, associé au Ca²⁺, active la protéine kinase C (PKC).
Résumé :
Récepteur → Gq/11 → PLCβ → PIP₂ → IP₃ + DAG → ↑ Ca²⁺ intracellulaire + activation de la PKC.
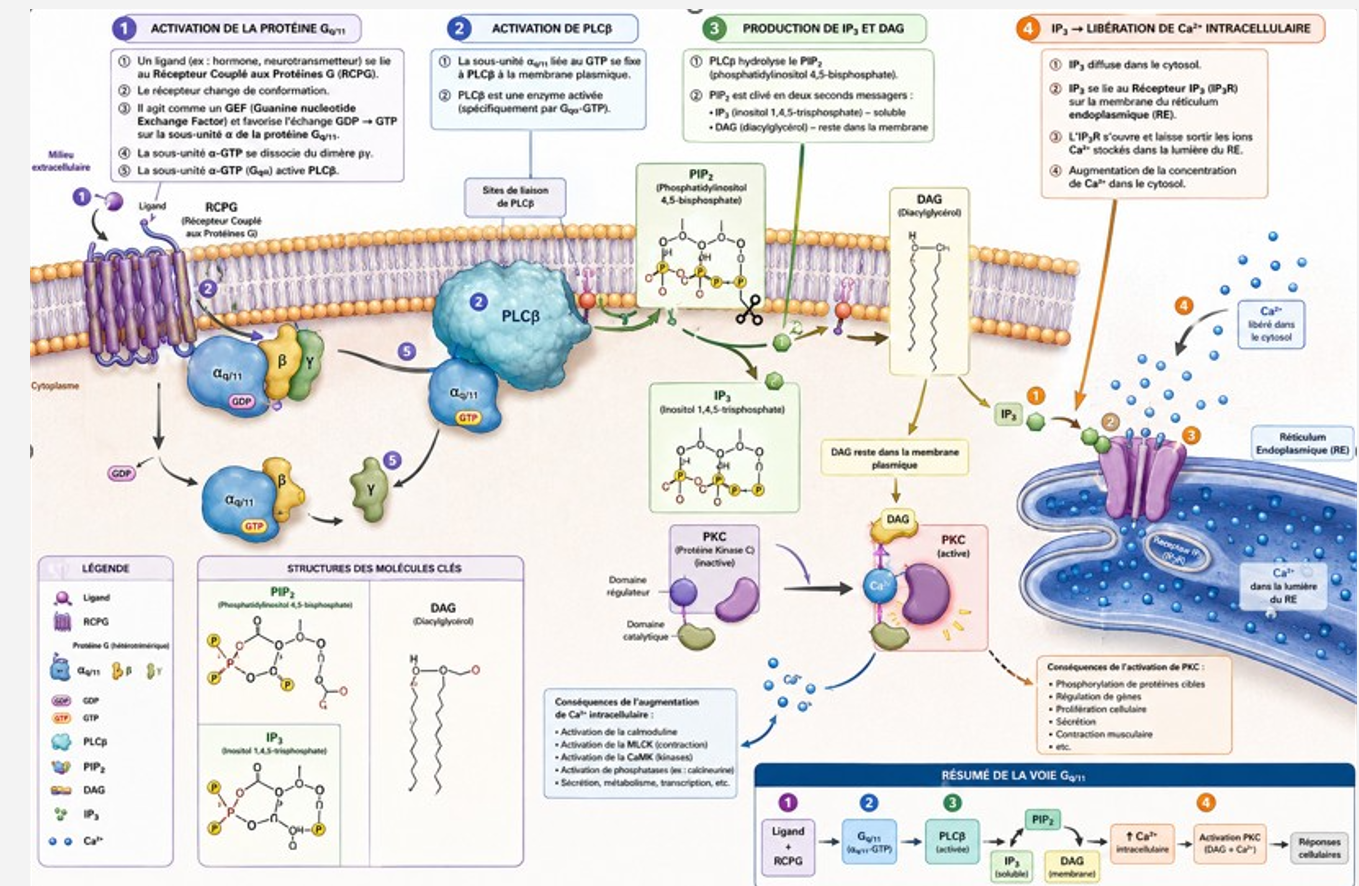
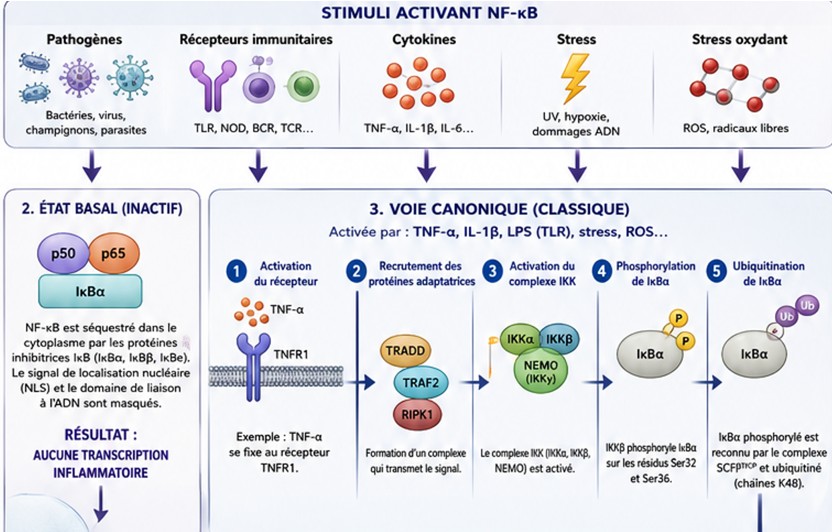
Voie 2 : Activation NF-κB
L’activation de NF-κB (Nuclear Factor kappa-light-chain-enhancer of activated B cells) constitue l’un des mécanismes moléculaires les plus importants de l’immunité innée, de l’inflammation, de la survie cellulaire et de nombreuses pathologies humaines. En biologie cellulaire moderne, NF-κB est souvent considéré comme le « maître régulateur » de la réponse inflammatoire.
NF-κB n’est pas une protéine unique mais une famille de facteurs de transcription.
Chez les mammifères, elle comprend cinq protéines :
| Protéine | Gène |
| RelA (p65) | RELA |
| RelB | RELB |
| c-Rel | REL |
| p50 | NFKB1 |
| p52 | NFKB2 |
Ces protéines possèdent un domaine commun : RHD (Rel Homology Domain) qui intervient dans la liaison à l’ADN, la dimérisation et l’interaction avec les inhibiteurs IκB
NF-κB agit sous forme de : homodimères, hétérodimères
Le plus fréquent est : p50/RelA (p65) qui est responsable de la majorité des réponses inflammatoires. État basal : NF-κB est inactif
Dans une cellule non stimulée :
NF-κB est retenu dans le cytoplasme. Il est lié à des protéines inhibitrices : IκBα, IκBβ et IκBε
Ces protéines masquent :
- le signal de localisation nucléaire (NLS)
- le domaine de liaison à l’ADN
Ainsi :
NF-κB + IκB
↓
Cytoplasme
↓
Aucune transcription inflammatoire
L’inflammation est donc maintenue sous contrôle.
Pourquoi activer NF-κB ?
L’organisme doit réagir rapidement à : infections (bactéries, virus, champignons et parasites), au stress (UV, radicaux libres, hypoxie et dommages de l’ADN) , aux cytokines (TNF-α, IL-1β) et aux Récepteurs immunitaires (TLR (Toll-Like Receptors), NOD receptors, BCR ,TCR
Toutes ces voies convergent vers NF-κB.
1.Voie canonique (classique)
La plus importante.
Activée par : (TNF-α, IL-1β, LPS bactérien, TLR, stress oxydant)
Étape 1 : Activation du récepteur
Exemple : TNF-α se fixe sur : TNFR1
Étape 2 : Recrutement des protéines adaptatrices
Formation d’un complexe :TRADD, TRAF2, RIPK1
Ces protéines transmettent le signal.
Étape 3 : Activation du complexe IKK
Le point clé.
Le complexe IKK contient :
| Sous-unité | Fonction |
| IKKα | kinase |
| IKKβ | kinase |
| NEMO (IKKγ) | régulatrice |
Signal inflammatoire
↓
Complexe IKK
Étape 4 : Phosphorylation de IκBα
IKKβ phosphoryle l’IκBα sur la Ser32 et Ser36
Étape 5 : Ubiquitination
La protéine phosphorylée est reconnue par : SCFβTrCP
qui ajoute :
- chaînes ubiquitine K48
IκBα
↓
Ubiquitination
Rôle du calcium
Le Ca²⁺ active, Calmoduline qui entraine l’activation de MLCK
Résultat final :
👉 Activation + augmentation de l’expression de MLCK
🔵 3. Fonction de MLCK
Réaction clé :
MLCK catalyse :
MLC + ATP → MLC phosphorylée (MLC-P)
Conséquence mécanique :
Sans stress :
- faible phosphorylation
- cytosquelette stable
- jonctions serrées intactes
Sous stress :
- ↑ phosphorylation de MLC
- contraction du cytosquelette actine-myosine
👉 Cela tire mécaniquement sur les jonctions serrées
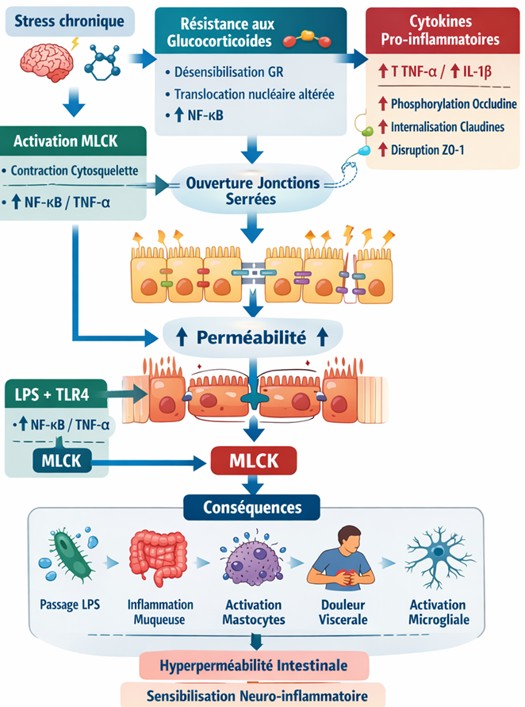
🔵 4. Désorganisation des tight junctions
Structure normale :
Protéines clés :
- Claudines
- Occludine
- ZO-1 / ZO-2
👉 Elles assurent l’étanchéité entre cellules
Après activation de MLCK :
4 effets majeurs :
- Contraction actomyosine
- Détachement de ZO-1
- Internalisation d’occludine
- Réorganisation des claudines
Résultat :
👉 Ouverture des espaces entre cellules
👉 ↑ perméabilité paracellulaire
🔵 5. Conséquences immunologiques
Une fois la barrière ouverte :
Passage de molécules :
- LPS (lipopolysaccharides)
- peptidoglycanes
- antigènes alimentaires
Activation immunitaire :
Les macrophages détectent ces éléments via :
👉 TLR4
Production de cytokines :
- TNF-α
- IL-6
- IL-1β
Conséquence :
👉 inflammation :
- locale (intestin)
- systémique (corps entier)
🔵 6. Boucle d’auto-entretien (cercle vicieux)
Le système s’auto-amplifie :
- Stress
- Activation mastocytes
- ↑ cytokines
- ↑ MLCK
- ↑ perméabilité
- entrée de toxines
- ↑ inflammation
👉 Retour au stress et aggravation
Ajout du microbiote :
- Dysbiose (déséquilibre bactérien)
- ↓ butyrate (protecteur intestinal)
👉 aggrave la situation
🔵 7. Régulation fine de MLCK
Régulation transcriptionnelle :
Facteurs activant MLCK :
- NF-κB
- AP-1
- C/EBPβ
Régulation post-traductionnelle :
- phosphorylation par kinases (PKC, MAPK)
- interaction avec cytosquelette
Modulateurs :
Inhibiteurs naturels :
- butyrate
- probiotiques
- AMPK
- antioxydants (N-acétylcystéine)
Activateurs :
- TNF-α
- LPS
- stress oxydatif
- dysbiose
🔵 8. Preuves expérimentales
Inhibition de MLCK :
- ML-7 (inhibiteur)
👉 restaure la barrière intestinale
Modèles génétiques :
- Knockout MLCK → résistance à l’hyperperméabilité
- Surexpression MLCK → perméabilité augmentée
Modèles de stress :
- ↑ MLCK
- ↑ MLC phosphorylée
- ↑ perméabilité
👉 effet bloqué si MLCK inhibée
🧩 Conclusion globale
MLCK agit comme :
👉 un interrupteur biomécanique
Il traduit :
- des signaux psychologiques (stress)
- en une réponse physique :
👉 ouverture de la barrière intestinale
🔥 Résumé ultra-synthétique
Stress → CRH + cytokines → ↑ Ca²⁺ → calmoduline → MLCK →
phosphorylation MLC → contraction → ouverture jonctions →
perméabilité → inflammation → cercle vicieux
💡 Ce que ça implique (important)
Ce mécanisme est impliqué dans :
- syndrome de l’intestin irritable (SII)
- maladies inflammatoires intestinales (MICI)
- troubles métaboliques
- troubles neuropsychiatriques (via axe intestin-cerveau)