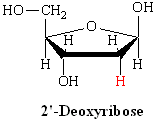
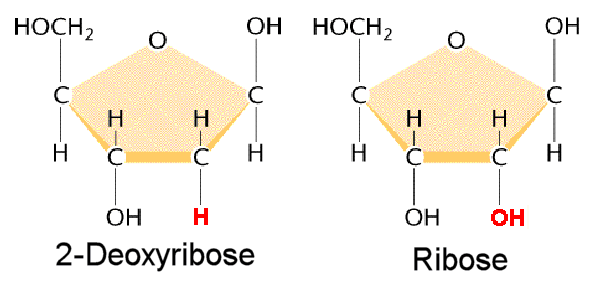
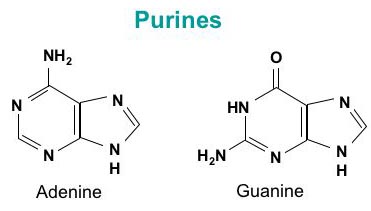
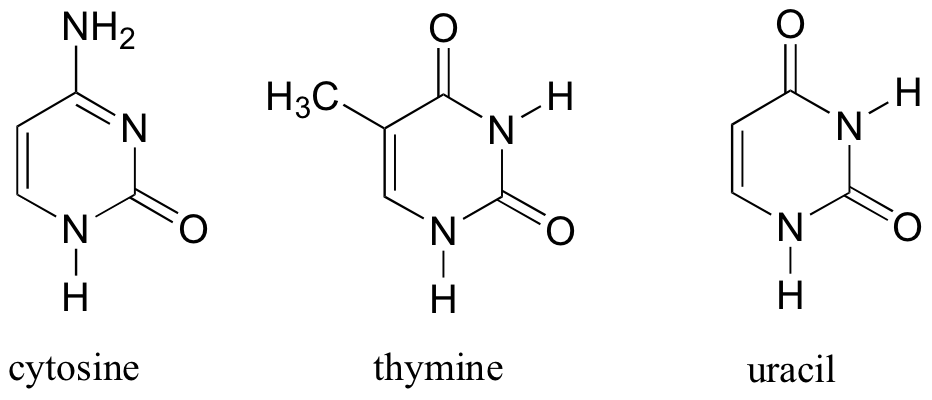
L’ADN est un polymère (une grande molécule qui contient des unités répétitives) composé d’un 2′ désoxyribose (un sucre à cinq carbones), d’un acide phosphorique et de 4 bases azotées notés A, T, G et C. Les structures chimiques des bases sont indiquées ci-dessous. Notez que deux des bases ont une structure à double anneau appelées purines. Les deux autres bases ont une structure à anneau unique, celles-ci sont appelées pyrimidines.
Les bases purines sont l’adénine (A) et guanine (G).
Les bases pyrimidiques sont la thymine (T) et la cytosine (c).
Dans l’ADN chaque base est liée à une molécule du sucre (désoxyribose) formant un composé appelé nucléoside.

Quand un groupe phosphate est également attaché à la molécule du sucre le nucléoside devient un nucléotide.

Ainsi un nucléotide est un nucléoside plus un phosphate. Dans la numérotation conventionnelle des atomes de carbone dans le sucre, l’atome de carbone auquel est attaché la base est le carbone 1′ (les atomes dans le sucre ont des numéros primes pour les distinguer des atomes dans les bases).
Dans les acides nucléiques, comme l’ADN et l’ARN, les nucléotides sont joints pour former une chaîne polynucléotidique dans laquelle le phosphate attaché à l’extrémité 5′ d’un carbone d’un sucre est lié au groupe hydroxyle fixé à l’extrémité 3′ de carbone du sucre suivant en ligne. Les liaisons chimiques par lesquelles les composants de sucre de nucléotides adjacents sont reliés par l’intermédiaire des groupes phosphate sont appelées liaisons phosphodiester.
L’orientation 5′-3′-5′-3 ‘de ces liens se poursuit tout au long de la chaîne formant un groupe 5′-phosphate (5′-P) à une extrémité et un groupe 3′-hydroxyle (3′-OH) à L’autre. L’asymétrie des extrémités d’un brin d’ADN crée une polarité dont le sens est déterminé par ces extrémités 5′ phosphate et 3′ hydroxyles.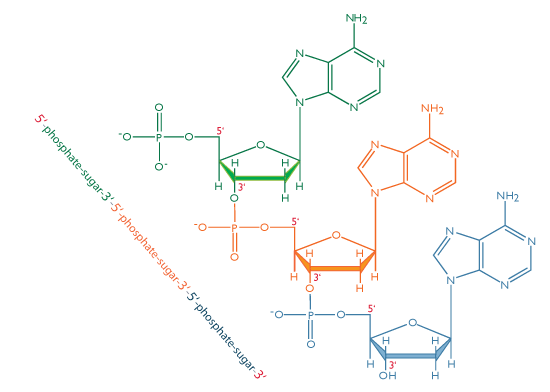
Trois ans avant la proposition de la structure tridimensionnelle essentiellement correcte de l’ADN par Watson et Crick comme une double hélice, Erwin Chargaff avait développé une technique chimique pour mesurer la quantité de chaque base présente dans l’ADN. Comme nous décrivons sa technique nous allons représenter la concentration molaire de chaque base par le symbole de la base entre crochets; par exemple, [A] désigne la concentration molaire de l’adénine. Chargaff a utilisé sa technique pour mesurer la [A], [T], [G] et [C] dans le contenu de l’ADN à partir d’une variété de sources. Il a constaté que dans la composition des bases de l’ADN, la quantité de G + C était constante dans toutes les cellules de l’organisme et à l’intérieur d’une espèce mais différait d’une espèce à l’autre.
Chargaff a également observé certaines relations régulières entre les concentrations molaires des différentes bases. Ces relations sont maintenant appelées les règles de chargaff:
La quantité d’adénine est égale à celle de thymine : [A] = [T].
La quantité de guanine est égale à celle de cytosine : [G] = [C].
La quantité des bases puriques est égale à celle des bases pyrimidiques : [A] + [G] = [T] + [C].
Bien que la base chimique de ces observations n’était pas connue à l’époque, l’une des caractéristiques intéressantes de la structure de Watson-Crick des brins complémentaires appariés était exactement ce qui était expliqué par les règles de Chargaff. Parce que A est toujours jumelé avec T dans l’ADN double brin il doit suivre ce que [A] = [T]. De même parce que G est appariée avec C, [G] = [C]. La troisième règle suit par addition des deux autres : [A] + [G] = [T] + [C].
La structure physique de la double hélice :
Dans la structure tridimensionnelle de la molécule d’ADN proposé en 1953 par Watson et Crick, la molécule est constituée de deux chaînes polynucléotidiques subissant une torsion,l’une autour de l’autre pour former une hélice double brin dans laquelle l’adénine, la thymine, la guanine et la cytosine sont appariées dans des brins opposés. Dans la structure standard, appelée la forme d’ADN B, chaque chaîne fait un tour complet tous les 34 A°.
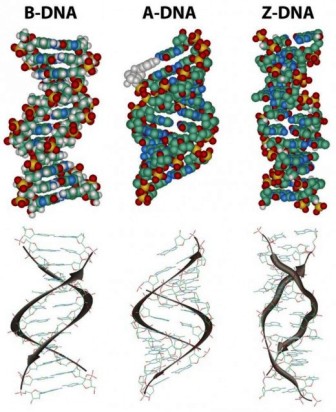
L’hélice est droitière ce qui signifie que, quand vous regardez le long de la structure de l’ADN, chaque chaîne suit un chemin dans le sens horaire à mesure qu’elle progresse. Les bases sont espacées à 3,4 A donc il y a dix bases par tour d’hélice dans chaque brin et dix paires de bases par tour de la double hélice. Chaque base est jumelée à une base complémentaire dans l’autre brin par des liaisons hydrogène, qui fournissent la principale force maintenant les brins ensemble (une liaison hydrogène est une liaison faible dans laquelle deux atomes chargés négativement partagent un atome d’hydrogène). Les paires de bases sont planes, parallèles entre elles et perpendiculaires à l’axe longitudinal de la double hélice.
Lors de l’examen d’une molécule d’ADN les biologistes moléculaires font souvent référence à des brins individuels, comme simples brins ou l’ADN simple brin et à double hélice comme l’ADN double brin ou à une molécule duplex. Les deux rainures en spirale à l’extérieur de la double hélice ne sont pas symétriques, une rainure, appelée le grand sillon, est plus grande que l’autre qui est appelée le sillon mineur. Les protéines qui interagissent avec l’ADN double brin ont souvent des régions qui entrent en contact avec les paires de bases en s’ajustant dans le grand sillon, dans le petit sillon ou dans les deux rainures.
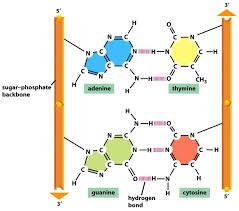
L’élément central de la structure de l’ADN est l’appariement des bases complémentaires A avec T et G avec C. Les liaisons hydrogène qui se forment dans la paire de bases adénine-thymine et dans la paire guanine-cytosine sont illustrées sur la figure ci-dessus notez qu’une paire A-T a deux liaisons hydrogène et qu’une paire de G-C a trois liaisons hydrogène. Cela signifie que la liaison hydrogène entre G et C est plus forte dans le sens où il faut plus d’énergie pour rompre, par exemple la quantité de chaleur nécessaire pour séparer les brins appariés dans une ADN duplex augmente avec le pour cent de G + C parce que rien ne limite la séquence de bases dans un seul brin, toute séquence peut être présente le long d’un brin. Ceci explique l’observation de Chargaff que l’ADN de différents organismes peut avoir différentes compositions de base. Cependant parce que les brins de l’ADN duplex sont complémentaires, règles de Chargaff de [A] = [T] et [G] = [C] sont vraies quelle que soit la composition des bases .
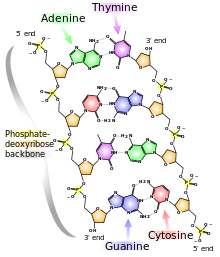
Chaque épine dorsale dans une double hélice se compose de sucres désoxyribose alternant avec des groupes phosphate qui lie l’atome de carbone 3′ d’un sucre à l’atome de carbone 5′ du sucre suivant sur la ligne. Les deux brins polynucléotidiques de la double hélice sont orientés dans des directions opposées dans le sens que les bases qui sont appariés sont liés aux sucres respectivement au-dessus et en-dessous du plan de couplage. Ce décalage existe parce que les liaisons phosphate dans les ossatures courent dans des directions opposées et les brins sont dits antiparallèles. Cela signifie que chaque extrémité de la double hélice possède un groupe 5′-P (sur un brin) et un groupe 3′-OH (de l’autre brin) comme le montre la figure ci-dessous :
Les diagrammes des ADN duplex sont statiques et donc quelque peu trompeurs. En réalité l’ADN est une molécule dynamique constamment en mouvement. Dans certaines régions les brins peuvent se séparer brièvement puis se réunir à nouveau dans la même conformation ou dans une autre. Bien que la double hélice droitière est la forme standard l’ADN peut constituer plus de 20 variantes légèrement différentes d’hélices droitières et dans certaines régions peut même exister des hélices dans lequeles les brins sont gauchers (appelées la forme Z de l’ADN) . S’ il y a des tronçons complémentaires de nucléotides dans le même brin et, si ce brin est séparé de son partenaire, il peut se replier sur lui-même comme une épingle à cheveux. Même les triples hélices composées de trois brins peuvent se former dans les régions de l’ADN qui contiennent des séquences de base appropriées.
Qu’est ce qu’un matériel a besoin pour jouer le rôle de transmission génétique ?
Chaque polymère ne serait pas utile comme matériel génétique. Cependant l’ADN est admirablement adaptée à une fonction génétique parce qu’elle satisfait aux trois exigences essentielles d’un matériel génétique. Premièrement, tout le matériel génétique doit pouvoir être reproduit avec précision de sorte que l’information qu’il contient soit précisément reproduit et hérité par les cellules fille. Dans l’ADN cela se fait au moyen d’un code génétique dans lequel des groupes de trois bases spécifient les acides aminés. Du fait que les quatre bases dans une molécule d’ADN peuvent être disposées dans un ordre quelconque et parce que la séquence peut varier d’une partie de la molécule à l’autre et d’un organisme à l’autre, l’ADN peut contenir un grand nombre de régions uniques, chacun d’eux pouvant être un gène distinct. Une longue chaîne d’ADN peut diriger la synthèse d’une grande variété de molécules de protéines.
Un matériel génétique doit aussi être capable de subir des mutations occasionnelles dans lesquelles l’information qu’il porte est modifiée. En outre les molécules mutantes doivent être capables de se répliquer aussi fidèlement que la molécule parentale de sorte que les mutations deviennent héréditaires. Watson et Crick ont suggéré que des mutations héréditaires pourraient être possibles dans l’ADN par mauvais appariement rare des bases avec le résultat qu’un nucléotide incorrect est incorporé dans un brin d’ADN en répliquation.