Chapter 2 : the chemical composition of DNA
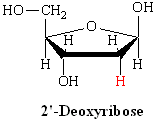
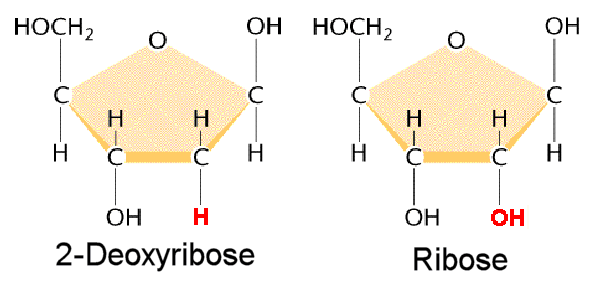
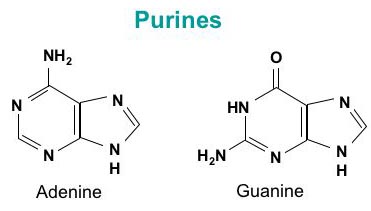
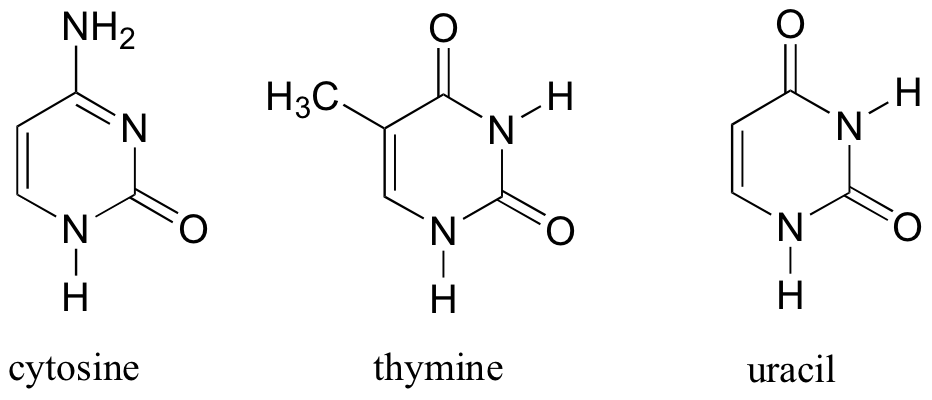
DNA is a polymer ( a large molecule that contains repeating units) composed of 2’ deoxyribose (a five-carbon sugar), phosphoric acid, and the four nitrogen containing bases denoted A, T, G and C. The chemical structures of the bases are shown below. Note that two of the bases have a double-ring structure, these are called purines. The other two bases have a single-ring structure, these are called pyrimidines.
The purine bases are adenine (A) and guanine (G).
The pyrimidine bases are thymine (T) and cytosine ©.
In DNA, each base is chemically linked to one molecule of the sugar deoxyribose, forming a compound called a nucleoside.

When a phosphate group is also attached to the sugar, the nucleoside becomes a nucleotide .

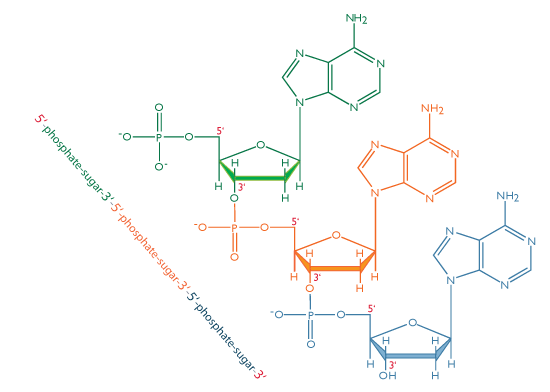
Thus a nucleotide is a nucleoside plus a phosphate. In the conventional numbering of the carbon atoms in the sugar,the carbon atom to which the base is attached is the 1’ carbon. (The atoms in the sugar are given primed numbers to distinguish them from atoms in the bases).
In nucleic acids, such as DNA and RNA, the nucleotides are joined to form a polynucleotide chain, in which the phosphate attached to the 5’ carbon of one sugar is linked to the hydroxyl group attached to the 3’ carbon of the next sugar in line . The chemical bonds by which the sugar components of adjacent nucleotides are linked through the phosphate groups are called phosphodiester bonds. The 5’-3’-5’-3’ orientation of these linkages continues throughout the chain are a 5’-phosphate (5’-P) group at one end and a 3’-hydroxyl (3’-OH) group at the other. The asymmetry of the ends of a DNA strand implies that each strand has a polarity determined by which end bears the 5’-phosphate and which end bears the 3’-hydroxyl.
Three years before Watson and Crick proposed their essentially correct three-dimensional structure of DNA as a double helix, Erwin Chargaff developed a chemical technique to measure the amount of each base present in DNA. As we describe his technique, we will let the molar concentration of any base be represented by the symbol for the base in square brackets; for example, [A] denotes the molar concentration of adenine. Chargaff used his technique to measure the [A], [T], [G], and [C] content of the DNA from a variety of sources. He found that the base composition of the DNA, defined as the present G+C, differs among species but is constant in all cells of an organism and within a species.
Chargaff also observed certain regular relationships among the molar concentrations of the different bases. These relationships are now called Chargaff’s rules:
The amount of adenine equals that of thymine: [A]=[T]
The amount of guanine equals that of cytosine: [G]=[C]
The amount of purine bases equals that of pyrimidine bases: [A]+[G]=[T]+[C]
Although the chemical basis of these observations was not known at the time, one of the appealing features of the Watson-Crick structure of paired complementary strands was that is explained Chargaff’s rules. Because A is always paired with T in double-stranded DNA, it must follow that [A]=[T]. Similarly, because G is paired with C, [G]=[C]. The third rule follows by addition of the other two: [A]+[G]=[T]+[C].
The Physical Structure of the Double Helix :
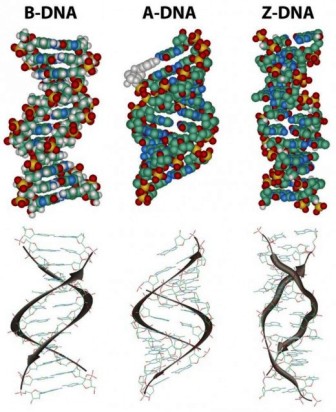
In the three-dimensional structure of the DNA molecule proposed in 1953 by Watson and Crick, the molecule consists of two polynucleotide chains twisted around one another to form a double-stranded helix in which adenine and thymine, and guanine and cytosine, are paired in opposite strands . In the standard structure, which is called the B form of DNA, each chain makes one complete turn every 34 A.
The helix is right-handed, which means that as you look down the barrel, each chain follows a clockwise path as it progresses. The bases are spaced at 3.4 A, so there are ten bases per helical turn in each strand and ten base pairs per turn of the double helix. Each base is paired to a complementary base in the other strand by hydrogen bonds, which provide the main force holding the strands together. (A hydrogen bond is a weak bond in which two negatively charged atoms share a hydrogen atom). The paired bases are planar, parallel to one another, and perpendicular to the long axis of the double helix. When discussing a DNA molecule, molecular biologists frequently refer to the individual strands as single strands or as single-stranded DNA; they refer to the double helix as double-stranded DNA or as a duplex molecule. The two grooves spiraling along outside of the double helix are not symmetrical; one groove, called the major groove, is larger than the other, which is called the minor groove. Proteins that interact with double-stranded DNA often have regions that make contact with the base pairs by fitting into the major groove, into the minor groove, or into both grooves.
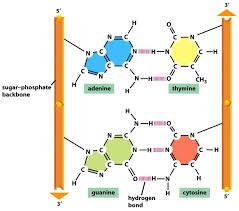
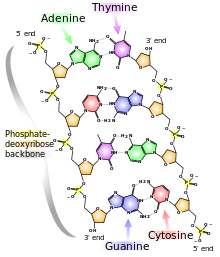
The central feature of DNA structure is the pairing of complementary bases, A with T and G with C. The hydrogen bonds that form in the adenine-thymine base pair and in the guanine-cytosine pair are illustrated in Figure above Note that an A-T pair has two hydrogen bonds and that a G-C pair has three hydrogen bonds. This means that the hydrogen bonding between G and C is stronger in the sense that it requires more energy to break; for example, the amount of heat required to separate the paired strands in a DNA duplex increases with the percent of G + C. Because nothing restricts the sequence of bases in a single strand, any sequence could be present along one strand. This explains Chargaff’s observation that DNA from different organisms may have different base compositions. However, because the strands in duplex DNA are complementary, Chargaff’s rules of [A] = [T] and [G] = [C] are true whatever the base composition.
Each backbone in a double helix consists of deoxyribose sugars alternating with phosphate groups that link the 3’ carbon atom of one sugar to the 5’ carbon of the next in line. The two polynucleotide strands of the double helix are oriented in opposite directions in the sense that the bases that are paired are attached to sugars above and below the plane of pairing, respectively. offset because the phosphate linkages in the backbones run in opposite directions , and the strands are said to be antiparallel. This means that each terminus of the double helix possesses one 5’-P group (on one strand) and one 3’-OH group (on the other strand), as shown in Figure below :
The diagrams of the DNA duplexes are static and so some-what misleading. DNA is a dynamic molecule, constantly in motion. In some regions, the strands can separate briefly and then come together again in the same conformation or in a different one. Although the right-handed double helix is the standard form, DNA can form more than 20 slightly different variants of right-handed helices, and some regions can even form helices in which the strands twist to the left (called the Z form of DNA). If there are complementary stretches of nucleotides in the same strand, then a single strand, separated from its partner, can fold back upon itself like a hairpin. Even triple helices consisting of three strands can form in regions of DNA that contain suitable base sequences.
What a Genetic Material Needs That DNA Supplies :
Not every polymer would be useful as genetic material. However, DNA is admirably suited to a genetic function because it satisfies the three essential requirements of a genetic material. First, any genetic material must be able to be replicated accurately, so that the information it contains is precisely replicated and inherited by daughter cells added to the end of a growing protein molecule. In DNA, this is done by means of a genetic code in which groups of three bases specify amino acids. Because the four bases in a DNA molecule can be arranged in any sequence, and because the sequence can vary from one part of the molecule to another and from organism to organism, DNA can contain a great many unique regions, each of which can be a distinct gene. A long DNA chain can direct the synthesis of a variety of different protein molecules.
A genetic material must also be capable of undergoing occasional mutations in which the information it carries is altered. Furthermore, the mutant molecules must be capable of being replicated as faithfully as the parental molecule, so that mutations are heritable. Watson and Crick suggested that heritable mutations might be possible in DNA by rare mispairing of the bases, with the result that an incorrect nucleotide becomes incorporated into a replicating DNA strand.
Chapter 1 : stem cells
What is a stem cell?
Ultimately, every cell in the human body can be traced back to a fertilized egg that came into existence from the union of egg and sperm. But the body is made up of over hundreds of different types of cells. All of these cell types come from a pool of stem cells in the early embryo. During early development, as well as later in life, various types of stem cells give rise to the specialized or differentiated cells that carry out the specific functions of the body, such as blood, muscle,bone and nerve cells.
Stem cells can regenerate themselves or produce specialized cell types. This property make stem cells appealing for scientists seeking to create medical treatments that replace lost or damaged cells.
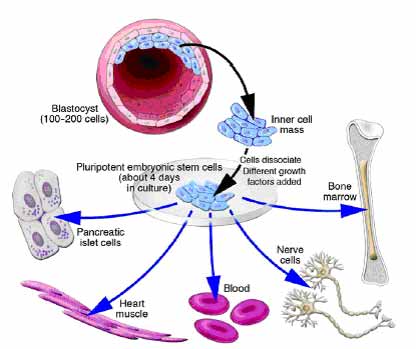
TYPES OF STEM CELLS :
Stem cells are found in all of us, from the early stages of human development to the end of life. All stem cells may prove useful for medical research, but each of the different types has both promise and limitations. Embryonic stem cells, which can be derived from a very early stage in human development, have the potential to produce all of the body’s cell type. Adult stem cells, which are found in certain tissues in fully developed humans, from babies to adults, may be limited to producing only certain types of specialized cells. Recently, scientists have also identified stem cells in umbilical cord blood and the placenta that can give rise to the various types of blood cells.
Embryonic Stem Cells :
A blastocyst, is a pre-implantation embryo that develops 5 days after the fertilization of an egg by a sperm. It contains all the material necessary for the development of a complete human being. The blastocyst is a mostly hollow sphere. In its interior is the inner cell mass, which is composed of 30-34 cells that are referred to by scientists as pluripotent because they can differentiate into all of the cell types of the body.
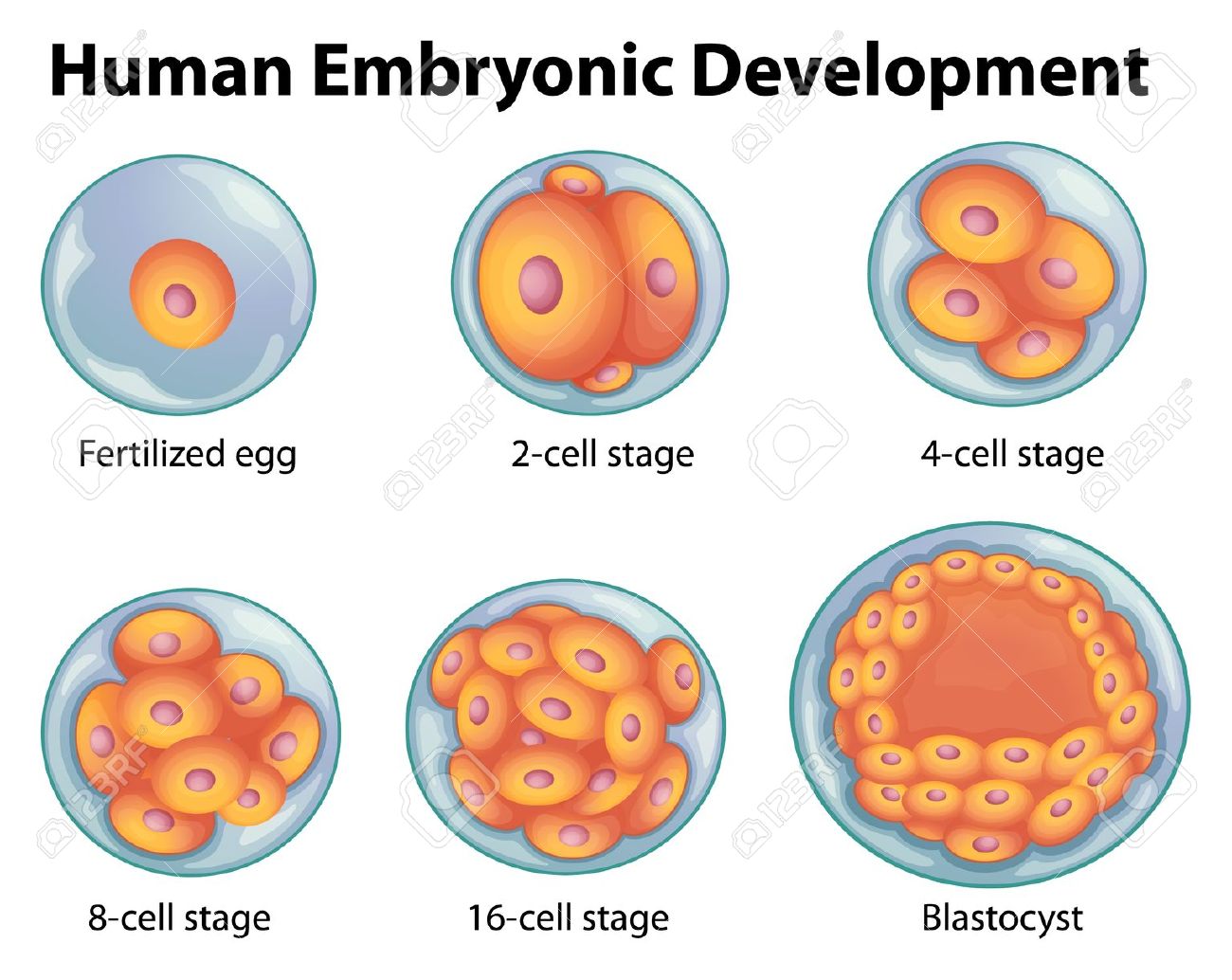
In normal development, the blastocyst would implant in the wall of the uterus to become the embryo and continue developing into a mature organism. Its outer cells would begin to from the placenta and the inner cell mass would begin to differentiate into the progressively more specialized cell types of the body.
When the blastocyst is used for stem cell research, scientists remove the inner cell masse and place these cells in a culture dish with a nutrient-rich liquid where they give rise to embryonic stem cells. Embryonic stem cells seem to be more flexible than stem cells found in adults, because they have the potential to produce every cell type in the human body. They are also generally easier to collect, purify and maintain in the laboratory than adult stem cells. Scientists can induce embryonic stem cells to replicate themselves in an undifferentiated state for very long periods of time before stimulating them to create specialized cells. This means that just a few embryonic stem cells can build a large bank of stem cells to be used in experiments. However, such undifferentiated stem cells could not be used directly for tissue transplants because they can cause a type of tumor called a teratoma. To be used for therapies, embryonic stem cells would first need to be differentiated into specialized cell types.
Some find embryonic stem cell research to be morally objectionable, because when scientists remove the inner cell mass, the blastocyst no longer has the potential to become a fully developed human being.
Sources of Embryonic Stem Cells (In Vitro Fertilization):
The largest potential source of blastocysts for stem cell research is from in vitro fertilization (IVF) clinics. The process of IVF requires the retrieval of a woman’s eggs via a surgical procedure after undergoing an intensive regimen of “fertility drugs,” which stimulate her ovaries to produce multiple mature eggs. When IVF is used for reproductive purposes, doctors typically fertilize all of the donated eggs in order to maximize their chance of producing a viable blastocyst that can be implanted in the womb. Because not all the fertilized eggs are implanted, this has resulted in a large bank of “excess” blastocysts that are currently stored in freezers around the country. The blastocysts stored in IVF clinic could prove to be a major source of embryonic stem cells for use in medical research. However, because most of these blastocysts were created before the advent of stem cell research, most donors were not asked for their permission to use these left-over blastocysts for research.
The in vitro fertilization (IVF) technique could potentially also be used to produce blastocysts specifically for research purposes. This would facilitate the isolation of stem cells with specific genetic traits necessary for the study of particular diseases. For example, it may be possible to study the origins of an inherited disease like cystic fibrosis using stem cells made from egg and sperm donors who have this disease. The creation of stem cells specifically for research using IVF is, however, ethically problematic for some people because it involves intentionally creating a blastocyst that will never develop into a human being.
Nuclear Transfer:
The process called nuclear transfer offers another potential way to produce embryonic stem cells. In animals, nuclear transfer has been accomplished by inserting the nucleus of an already differentiated adult cell-for example, a skin cell-into a donated egg that has had its nucleus removed. This egg, which now contains the genetic material of the skin cell, is then stimulated to form a blastocyst from which embryonic stem cells can be derived. The stem cells that 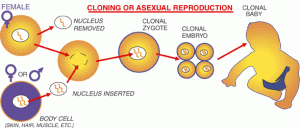 are created in this way are therefore copies or “clones” of the original adult cell because their nuclear DNA matches that of the adult cell.
are created in this way are therefore copies or “clones” of the original adult cell because their nuclear DNA matches that of the adult cell.
As of the summer of 2006, nuclear transfer has not been successful in the production of human embryonic stem cells, but progress in research suggests that scientists may be able to use this technique to develop human stem cells in the future.
Producing Embryonic Stem Cells Using Nuclear Transfer Is Not the Same as Reproductive Cloning
The use of nuclear transfer to develop disease-specific stem cells can be called research cloning, and the use of this technique for personalized tissue transplants is sometimes called therapeutic cloning. These terms must be carefully distinguished from reproductive cloning, in which the intent is to implant a cloned embryo in a female’s womb and allow it to develop fully into an individual. This was the technique by which Dolly the sheep was made and is now widely used for reproductive cloning in animals. In humans, however, reproductive cloning has been actively discouraged by most in the scientific community.
Scientists believe that if they are able to use nuclear transfer to derive human stem cells, it could allow them to study the development and progression of specific diseases by creating stem cells containing the genes responsible for certain disorder. In the future, scientists may also be able to create “personalized” stem cells that contain only the DNA of a specific patient. The embryonic stem cells created by nuclear transfer would be genetically matched to a person needing a transplant, making it far less likely that the patient’s body would reject the new cells than it would be with traditional tissue transplant procedures.
Although using nuclear transfer to produce stem cells is not the same as reproductive cloning, some are concerned about the potential misapplication of the technique for reproductive cloning purposes. Other ethical considerations include egg donation, which requires informed consent, and possible destruction of blastocysts.
Adult Stem Cells :
Adult stem cells are hidden deep within organs, surrounded by millions of ordinary cells, and may help replenish some of the body’s cells when needed. In fact, some adult stem cells are currently being used in therapies. They have been found in several organs that need a constant supply of cells, such as the blood, skin and lining of the gut, and have also been found in surprising places like the brain, which is not known to readily replenish its cells. Unlike embryonic stem cells, adult stem cells are already somewhat specialized. For example, blood stem cells normally only give rise to the many types of blood cells, and nerve stem cells can only make the various types of brain cells. Recent research however, suggests that some adult stem cells might be more flexible than previously thought, and may be made to produce a wider variety of cells types. For example, some experiments have suggested that blood stem cells isolated from adult mice may also be able to produce liver, muscle and skin cells, but these results are not yet proven and have not been demonstrated with human cells. Nevertheless, scientists are working on finding a way to stimulate adult stem cells, or even other types of adult cells, to be more versatile. If they succeed, it could provide another source of unspecialized stem cells.
Identifying Stem Cells :
As early as 1961, scientists knew that adult bone marrow contained cells that could make all of the blood cell types. But it wasn’t until 1988 that those stem cells were isolated as pure populations. Why did it take so long? The techniques for identifying stem cells have only recently been developed. Partly, this is because adult stem cells are, by their very nature, inconspicuous in shape, size and function. They also tend to hide deep in tissues and are present only in very low numbers, making their identification and isolation like finding a needle in a haystack.
How do scientists know when they have found a stem cell? Every cell displays an array of proteins on its surface; different cell types have different proteins. Scientists can use these surface proteins as “markers” that characterize individual cell types – a type of molecular ID. For example, using molecules that recognize and attach to specific surface proteins and that can fluoresce under certain wavelengths of light, scientists can visually tell the difference between a blood cell. Unfortunately, not all stem cells can now be identified in this manner because scientists have not yet identified markers for all stem cell types. Scientists also identify stem cells by observing their behavior in the laboratory: stem cells must be able to remain unspecialized and self-renew for long periods of time.
Scientists believe that there might be more types of adult stem cells than the handful that have already been identified, but finding them is a difficult process.
Culturing Cell Lines and Stimulating Stem Cells to Differentiate
Cell culture is a term that refers to the growth and maintenance of cells in a controlled environment out-side of an organism. A successful stem cell culture is one that keeps the cells healthy, dividing and unspecialized. The culturing of stem cells in the first step in establishing a stem cell line – a propagating collection of genetically identical cells. Cell lines are important because they provide a long-term supply of multiplying cells that can be shared among scientists for research and therapy development. The National Academies report Stem Cells and the Future of Regenerative Medicine (2001) described some of the challenges of maintaining cell lines: “Over time, all cell lines…change, typically accumulating harmful genetic mutations. There is no reason to expect stem cell lines to behave differently. While there is much that can be learned using existing stem cell lines…such concerns necessitate continued monitoring of these cells as well as the development of new stem cell lines in the future.”
Once they have established a stable stem cell line, scientists start the process of causing the stem cells to differentiate into specialized cell types. The cellular environment in which stem cells naturally reside provides scientists with clues about how to make them differentiate in a culture dish. For example, in the bone marrow, where blood stem cells reside, bone cells send physical and chemicals signals that tell the blood stem cells when to differentiate. Scientists are just beginning to understand these signals and have developed ways to mimic the natural processes in cell cultures. Usually, the technology involves adding certain proteins to the cell culture and, in some cases, introducing specific genes into the stem cells.
It will be essential that scientists are sure that stem cells have fully differentiated before they can use them for medical applications. If completely undifferentiated stem cells (such as embryonic stem cells) are implanted directly into an organism, they can cause a type of tumor called a teratoma, which scientists have observed in experiments using mice. Semi-specialized adult stem cells and differentiated cells derived from embryonic stem cells are unlikely to cause teratomas.
Alternatives to Using Embryos in Stem Cell Research
To address ethical concerns about the destruction of blastocysts, scientists are trying to find new ways of obtaining stem cells that behave like embryonic stem cells but that don’t require harming a blastocyst. As the science progresses, ethical issues surrounding these alternatives may also arise. Some possible alternatives include:
1) Cells collected from the morul, the developmental stage prior to the blastocyst. The morula, a solid ball of about 16-30 cells, seems able to sustain the loss of a few cells without developmental damage so that the remaining cells can continue to develop. Cell extraction from the morula is already being used in some clinics to screen for genetic disorders in embryos produced by in vitro fertilization. Researchers have recently shown that cells isolated from a mouse morula can give rise to embryonic stem cells while the remaining morula cells develop into a healthy mouse. However, this process may still be morally objectionable to some because of the chance of harm to the morula, and because the long-term effects of removing cells from a morula are not yet known.
2) The creation of embryonic stem cells through a process called altered nuclear transfer (ANT). In this variation of the nuclear transfer technique, scientists create a blastocyst create whose genetic material has been changed so that further development and implantation into the uterus is not possible. It aims to create embryo-like entities that are not truly embryos but that can be a source of pluripotent stem cells. ANT, so far only tested with mouse blastocysts, could allow the creation of embryonic stem cells without destroying a viable human blastocyst. Some who object to embryonic stem cell research support ANT because the resulting blastocyst could never develop into a full human being and therefore would not have the moral status of a human embryo. However, this procedure is objectionable to some because they believe that it involves the creation of an imperfect blastocyst that is designed to be destroyed.
3) Causing an adult cell to act like an embryonic stem cell. During development, as cells become more and more specialized, they gradually lose the ability to turn on the genes that allow embryonic stem cells to be so versatile. The silencing if these genes seems to be responsible for keeping specialized cells specialized and limiting the differentiation capacities of adult stem cells. By “reprogramming” adult stem cells so that they can turn on the genes that allow versatility, scientists hope to cause them to revert to a more flexible state. It is even possible that scientists could one day “reprogram” any cell, not only stem cells. However, research in this area is in the early stages and scientists may be many years away from making an adult cell as versatile as an embryonic stem cell.